Monograph |
|
Corresponding author: Léo-Paul M. J. Dagallier ( leopauldagallier@gmail.com ) Academic editor: Andrés E. Ortiz-Rodriguez
© 2023 Léo-Paul M. J. Dagallier, Frank M. Mbago, Marie Couderc, Myriam Gaudeul, Aurélie Grall, Caroline Loup, Jan J. Wieringa, Bonaventure Sonké, Thomas L. P. Couvreur.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Dagallier L-PMJ, Mbago FM, Couderc M, Gaudeul M, Grall A, Loup C, Wieringa JJ, Sonké B, Couvreur TLP (2023) Phylogenomic inference of the African tribe Monodoreae (Annonaceae) and taxonomic revision of Dennettia, Uvariodendron and Uvariopsis. PhytoKeys 233: 1-200. https://doi.org/10.3897/phytokeys.233.103096
|
Abstract
Monodoreae (Annonaceae) is a tribe composed of 11 genera and 90 species restricted to the tropical African rain forests. All the genera are taxonomically well circumscribed except the species rich genera Uvariodendron and Uvariopsis which lack a recent taxonomic revision. Here, we used a robust phylogenomic approach, including all the 90 currently accepted species, with several specimens per species, and based on more than 300 Annonaceae-specific nuclear genes, to infer the phylogenetic tree of the Monodoreae and test the limits between the genera and species. We recover all the genera as monophyletic, except the genus Uvariopsis for which the species Uvariopsis tripetala falls outside this clade. We thus reinstate the monotypic genus Dennettia for its single species Dennettia tripetala. We also erect a new tribe, Ophrypetaleae trib. nov., to accommodate the genera Ophrypetalum and Sanrafaelia, as we recover them excluded from the Monodoreae tribe with good support. Below the genus level, the genera Isolona, Monodora, Uvariastrum, Uvariodendron and Uvariopsis show weakly supported nodes and phylogenetic conflicts, suggesting that population level processes of evolution might occur in these clades. Our results also support, at the molecular level, the description of several new species of Uvariodendron and Uvariopsis, as well as several new synonymies. Finally, we present a taxonomic revision of the genera Dennettia, Uvariodendron and Uvariopsis, which contain one, 18 and 17 species respectively. We provide a key to the 11 genera of the Monodoraeae and describe four new species to science: Uvariodendron kimbozaense Dagallier & Couvreur, sp. nov., Uvariodendron mossambicense Robson ex Dagallier & Couvreur, sp. nov., Uvariodendron pilosicarpum Dagallier & Couvreur, sp. nov. and Uvariopsis oligocarpa Dagallier & Couvreur, sp. nov., and provide provisional descriptions of three putatively new species. We also present lectotypifications and nomenclatural changes implying synonymies and new combinations (Uvariodendron citriodorum (Le Thomas) Dagallier & Couvreur, comb. et stat. nov., Uvariodendron fuscum var. magnificum (Verdc.) Dagallier & Couvreur, comb. et stat. nov., Uvariopsis congensis var. angustifolia Dagallier & Couvreur, var. nov., Uvariopsis guineensis var. globiflora (Keay) Dagallier & Couvreur, comb. et stat. nov., and Uvariopsis solheidii var. letestui (Pellegr.) Dagallier & Couvreur, comb. et stat. nov.).
Résumé
La tribu des Monodoreae (Annonaceae) est composée de 11 genres et 90 espèces des forêts tropicales humides d’Afrique. Tout les genres sont taxonomiquement bien résolus, à part les genres Uvariodendron et Uvariopsis qui manquent d’une révision taxonomique récente. Ici, nous avons utilisé une approche phylogénomique robuste pour estimer l’arbre phylogénétique des Monodoreae, et tester les limites de genres et d’espèces. Pour cela, nous avons inclut les 90 espèces acceptées, et avons séquencé plus de 300 gènes. Tous les genres sont retrouvés monophylétiques, à part le genre Uvariopsis pour lequel l’espèce Uvariopsis tripetala se retrouve exclue. Nous rétablissons donc le genre monotypique Dennettia et son unique espèce Dennettia tripetala. Nous érigeons une nouvelle tribu, les Ophrypetaleae trib. nov., pour accueillir les genres Ophrypetalum et Sanrafaelia, car nous les retrouvons exclus de la tribu des Monodoreae avec un bon support. Au niveau infra-générique, les genres Isolona, Monodora, Uvariastrum, Uvariodendron et Uvariopsis montrent de faibles supports de noeuds et des conflits phylogénétiques, ce qui suggère que des processus d’évolution se déroulent au niveau des populations. Nos résultats soutiennent également, sur un plan moléculaire, la description de plusieurs nouvelles espèces d’Uvariodendron et d’Uvariopsis, de même que plusieurs synonymies. Enfin, nous présentons une révision taxonomique des genres Dennettia, Uvariodendron et Uvariopsis, qui contiennent respectivement un, 18 et 17 espèces. Nous fournissons une clé des 11 genres de Monodoreae, et décrivons quatre nouvelles espèces pour la science: Uvariodendron kimbozaense Dagallier & Couvreur, sp. nov., Uvariodendron mossambicense Robson ex Dagallier & Couvreur, sp. nov., Uvariodendron pilosicarpum Dagallier & Couvreur, sp. nov. et Uvariopsis oligocarpa Dagallier & Couvreur, sp. nov., et fournissons une description provisoire de trois autres potentielles. Nous effectuons des lectotypifications et des changements nomenclaturaux tels que des synonymies et des nouvelles combinaisons (Uvariodendron citriodorum (Le Thomas) Dagallier & Couvreur, comb. et stat. nov., Uvariodendron fuscum var. magnificum (Verdc.) Dagallier & Couvreur, comb. et stat. nov., Uvariopsis congensis var. angustifolia Dagallier & Couvreur, var. nov., Uvariopsis guineensis var. globiflora (Keay) Dagallier & Couvreur, comb. stat. nov., et Uvariopsis solheidii var. letestui (Pellegr.) Dagallier & Couvreur, comb. stat. nov.).
Key words
conservation, evolution, new species, phylogeny, tropical rain forests
Introduction
Annonaceae is a diverse pantropical family of trees, shrubs and lianas mostly restricted to tropical rain forests. Recent work based on molecular phylogenetics has classified Annonaceae into four subfamilies and several tribes (
Monodoreae is a diverse clade of trees and small shrubs with one species (Monodora crispata) reported to be a lianescent (
In contrast to the recent taxonomic revisions cited above, recent revisionary work for the two most diverse genera of the tribe, Uvariodendron and Uvariopsis, are more than 80 years old (
Reconstruction of phylogenetic relationships in species-rich clades such as Annonaceae is challenging. Several phylogenetic analyses have focused on the whole family (
Species level relationships within Monodoreae have only been inferred, based on few chloroplast markers, in Isolona, Monodora and more recently in Mischogyne (
The aim of the present study is to generate a robust species-level phylogenomic tree of the Monodoreae and test the limits between the genera and species. Given that the species rich genera Uvariodendron and Uvariopsis have never been fully assessed phylogenetically and taxonomically, we give a particular focus to these two genera and present a taxonomic revision of these two genera. We discuss species delimitation based on both molecular and morphological evidence.
Materials and methods
Taxonomic revision
We examined the morphological characters of the Monodoreae species based on the protologues of the species and on the taxonomic revisions of those genera that were available (
When a holotype was composed of several sheets from a same gathering within a herbarium, we designated a single sheet as holotype (identified by its barcode) with the mention ‘sheet here designated’. The other sheets were listed as isotypes. This was done for all names except when the protologue explicitely specified that the holotype is composed of several sheets from a same gathering within a herbarium (e.g. Uvariodendron fuscum var. magnificum),
Measurements, colors and other details given in the descriptions are based on living material, spirit and herbarium specimens, and data derived from field notes. Morphological descriptions followed terminologies from
For this work, we applied a combination of the phylogenetic and morphological species concept. Thus, a species is recognized if it has support for monophyly and if it exhibits morphological characters that can differentiate it from the others. We also recognized varieties (or sunk previously described species into varieties) when a difference of morphology could be identified, but no monophyly was recovered by the phylogenetic analyses. In some cases, we recognized varieties even if they present an overlap in some of the morphological characters used to differentiate them. This is to recognize the existence of different morphological groups (or “archetypes”) in the morphological variation of the species. This reflects the uncertainty around the species, given the material available. The collection of additional material will certainly provide a better understanding of these morphologically complex groups.
To avoid ambiguities, when necessary, the names Uvariodendron and Uvariopsis were abbreviated to Ud. and Up., respectively.
IUCN assessments were retrieved from https://www.iucnredlist.org/ (accessed 21 September 2022). For species not present in the Red List, we made a preliminary conservation status assessment. We calculated the extent of occurrence (EOO) and the area of occupancy (AOO) using the ConR package (
Phylogenomics
Taxon sampling
We sampled at least one specimen for every species of the Monodoreae tribe, several specimens from the Uvarieae tribe representing different genera, as well as several other Annonaceae outgroups (
For all specimens, DNA was extracted from either silicagel dried leaf samples or leaf material taken from herbarium specimens. Details about specimens and vouchers are available in Suppl. material
For the downstream analyses, every sample was assigned a unique identifier as follow: RUN_##_I##_T##_Genus_species-COL_NUM. “RUN_##_I##_T##” is the identifier used for the sequencing process and bioinformatics analyses. “Genus_species” is the most up to date taxonomic identification of the specimen (i.e. after applying the taxonomic changes treated in this paper). To ease the reading of updated names, some of the specimens have their old identification in parentheses like “Genus_species_(ex_old name)”. “COL_NUM” is the three first letters of the name of the collector and the number of collection of the specimen (e.g. “VAL_2540” stands for the specimen n°2540 collected by J.L.C.H van Valkenburg). The details about every sample can be found in the Suppl. material
DNA extraction, library preparation and sequencing
DNA extraction and NGS library preparations for nuclear exon capture followed
Raw sequence bioinformatics
Raw read sequences were demultiplexed using the demultadapt script (https://github.com/Maillol/demultadapt). Adapters were removed from the reads using cutadapt 1.18 (
Contig assembly and alignment
To process the clean reads, we used the HybPiper v1.3.1 pipeline (
Phylogenetic reconstruction
We used two different approaches for the phylogenetic reconstruction: the concatenation approach and the gene tree approach. For each approach, we first removed the supercontigs that were putatively identified as paralogs within the HybPiper pipeline, and then filtered our dataset to select only the supercontigs that are present in 75% of the individuals and for which the sequence contains at least 75% of the length of the target sequence (75/75 filter, following
Concatenation approach
For each supercontig, the alignments were filled with gaps and missing individuals so that every sequence in the alignment has the same length and every alignment contains all the individuals. The alignments were then concatenated into a single “supermatrix” using the pxcat function in the phyx program (
Gene trees approach
We inferred a phylogenetic tree for every supercontig using RAxML 8.2.9 (
Results
Specimens examined
In total, we examined ca. 1,500 herbarium sheets, representing 928 herbarium specimens, including 486 of Uvariodendron, 405 of Uvariopsis, 35 of Dennettia, seven of Monocyclanthus and two of Lukea (Suppl. material
Morphological comparison of the Monodoreae genera. In bold, the combination of morphological characters diagnostic to each specific genus. H:w, height:width ratio.
| Number of accepted species | Leaf length | Tertiary veination | Sepals | Petals | Receptacle H:w | Flower sexuality | Carpels | |
|---|---|---|---|---|---|---|---|---|
| Asteranthe | 2 | 40–165 (180) mm | reticulate | 3, free to shortly connate at base | 2 whorls of 3, fused at base, inner and outer petals subequal | ca. 1 | bisexual | free |
| Dennettia | 1 | < 150 mm long | reticulate | 3, fused at base, forming a persistent ring with the 3 lobes generally clearly distinct, curved downward on the fruiting pedicel | 1 whorl of 3, free | < 1 | bisexual | free |
| Hexalobus | 5 | 36–360 mm long | reticulate | 3, free to shortly connate at base | 1 whorl of 6, fused at base, folded in bud | ca. 1 | bisexual | free |
| Isolona | 20 | 50–280 mm long | between reticulate and percurrent | 3, free, valvate | 1 whorl of 6, fused at base | < 1 | bisexual | fused (true syncarpy) |
| Lukea | 2 | 68–210 mm long | prominent reticulate | 3, fused, forming a bowl-shaped receptacle, persistent on the fruiting pedicel | 1 whorl of 3, free | > 1 | bisexual | free |
| Mischogyne | 5 | 70–320 mm long | prominent reticulate | 3, free, reduplicate-valvate in bud | 2 whorls of 3, free | > 1 | bisexual | free, connivent at base and separated and slightly curved outwards at apex |
| Monocyclanthus | 1 | > 150 mm long | reticulate | 3, fused at base, forming a ring with the lobes generally unconspicious, persistent on the fruiting pedicel | 1 whorl of 6, free | > 1 | bisexual | free |
| Monodora | 14 | 40–500 mm long | reticulate or more rarely percurrent | 3, free, valavate | 2 whorls of 3, fused at base, outer petals longer than inner petals | <1 to 1 | bisexual | fused (true syncarpy) |
| Uvariastrum | 5 | 60–220 mm long | reticulate | 3, free, reduplicate-valvate in bud, enclosing the petals in bud | 2 whorls of 3, free | ca. 1 | bisexual | free, connivent all along their length |
| Uvariodendron | 18 | 65–750 mm long | reticulate | 3, connivent to fused at base | 2 whorls of 3, free, inner and outer subequal in length, inner petals connivent at apex | ca. 1 | bisexual | free |
| Uvariopsis | 17 | 70–615 mm long | reticulate | 2, free to fused at base | 1 whorl of 4 (1 whorl of 3 in Uvariopsis congolana), free to fused | 1–1.2 | unisexual (except for Uvariopsis bisexualis) | free |
Phylogenomics
A total of 464 exon regions were recovered out of the 469 contained by the Annonaceae baiting kit (
Phylogenetic relationships
At the genus level and above, we recovered the same topology with both the gene tree (ASTRAL) and the concatenation (RAxML) approaches (Fig.

Phylogenetic trees of the Monodoreae inferred with ASTRAL, based on 334 nuclear genes trees. The branch support is given as local posterior probability (LPP) in three shades of greys. For the details on the specimens see Suppl. material
Species monophyly within the genera Asteranthe, Hexalobus, Lukea, Mischogyne, and Monocyclanthus received strong support, and the relationships between species were generally strong (BS > 90%, LPP > 0.9) and similar between both phylogenetic approaches (Fig.
In Isolona, Monodora, Uvariastrum, Uvariodendron and Uvariopsis, there are incongruences in topologies between the RAxML and the ASTRAL trees, and the node support is generally strong (BS > 90%, LPP > 0.9), but some nodes are weakly (BS < 70%, LPP < 0.7) to moderately supported (70% < BS < 90%, 0.7 < LPP < 0.9) (Fig.
In Uvariastrum, U. pierreanum and U. insculptum are strongly supported as monophyletic, but U. zenkeri is recovered as paraphyletic. The species U. germainii is retrieved as sister to U. pierreanum with moderate to strong support in both approaches. U. insculptum is retrieved as sister to all the other species in the genus (Fig.
In Uvariodendron and Uvariopsis, most species were recovered as monophyletic (Fig.
In Uvariopsis, the crown node of the clade formed by Up. bakeriana, Up. citrata, Up. korupensis, Up. submontana (« large leaves » group on Fig.
Discussion
Phylogenomics of the Monodoreae tribe
We provide here (Fig.
Generic level relationships within Monodoreae generally agree with molecular phylogenies based on few plastid markers (
Even though sequencing of hundreds of markers combined with a dense taxon sampling appear as a silver bullet to understand species limits, it doesn’t always provide clear cut answers. Indeed, phylogenetic conflicts are common in plant systematics: incongruences can occur between concatenation and gene tree approaches (
Re-circumscription of the tribe
The two monotypic sister genera from East Africa, Ophrypetalum Diels (Kenya and Tanzania) and Sanrafaelia Verdc. (Tanzania) (
The Ophrypetalum–Sanrafaelia clade share several morphological characters also found in other Monodoreae genera (Fig.

Ophrypetaleae tribe. Ophrypetalum odoratum Diels A trunk B young branch with leaves, upper side C flower and flower buds D flower, from below. Sanrafaelia ruffonammari Verdc E trunk F young branch with leaves and flower buds G flower buds H flower, top view I flower, side view J fruit. A, B Dagallier 54 C Couvreur 56 D Couvreur 82 E–J Dagallier 31. Photos A,,B, E–J Léo-Paul Dagallier C, ,D Thomas Couvreur.
After our study presented here, the tribe Monodoreae is composed of 11 genera and 90 species, and the main morphological characters are provide in Table
Asteranthe
Asteranthe is an East African genus characterised by large fleshy flowers with basally fused petals and numerous free carpels (
In addition,
Hexalobus
The genus Hexalobus contains five accepted species (
The widespread central African species H. crispiflorus is recovered as sister to the more narrowly distributed H. salicifolius (Cameroon and Gabon) with maximum support in both analyses (Fig.
Uvariastrum
Uvariastrum is a genus containing five accepted species characterized by reduplicate-valvate (i.e. curved outwards) sepals margins (
The case of the newly described genus Lukea
Recently, the genus Lukea Cheek & Gosline was described, endemic to East Africa with two species: Lukea quentinii Cheek & Gosline and Lukea triciae Cheek & Gosline (
Mischogyne
Mischogyne is a genus of five accepted species characterized by the combination of different morphological features such as prominent reticulate tertiary veins and carpels touching each other at their base but separated at their apex (
Uvariopsis
Uvariopsis Engl. is one of the most diverse genera of the Monodoreae tribe with 17 currently accepted species and until recently, several species were unknown to science (
Dennettia, Monocyclanthus and flower variations in Uvariopsis
In 1913, Baker described the monotypic genus Dennettia Baker f., with a single species Dennettia tripetala Baker f. having bisexual flowers with “2 rarely 3 sepals” and three petals in a single whorl (
After careful examination of 16 flowering specimens identified as Uvariopsis tripetala (including the type Dennett 44), we found that this species invariably exhibits 3 sepals and 3 petals. The sepals are fused at the base up to more than half of their length, forming a ring curved downward and persisting on the fruiting pedicel. In some rare cases we observed that the lobes of the calyx (i.e. the free parts at the apices of the basally fused sepals) are sometimes almost inconspicuous on the fruiting pedicel, allowing for the confusion with a two sepal-like calyx. To date, and based on the specimens we observed, we haven’t seen any flowers of Up. tripetala with four petals, contrary to the affirmation of
All these morphological and phylogenetic elements suggest that Dennettia (Uvariopsis) tripetala is part of neither the genus Uvariopsis nor the genus Monocyclanthus. We thus reinstate the monotypic genus Dennettia, with its single species Dennettia tripetala Baker f. (see Taxonomic treatment). This was also suggested previously by
Given the above, the morphological circumscription of Uvariopsis must be reevaluated. Uvariopsis was generally circumscribed as having 2 sepals and 4 petals (
The “large leaves” group
Based on our dense taxon sampling, we identified a group of closely related species for which the phylogenetic limits remain uncertain. Indeed, the four species Up. bakeriana, Up. citrata, Up. korupensis and Up. submontana clustered as monophyletic with strong support (Fig.
Based solely on the phylogeny, this group could be considered as a single species. However, given the morphological differences of each species detailed above we choose to keep the taxonomy of this group as it is. Phylogenetic conflict is known to be coincident with morphological innovations (
New and under collected species
In addition to the results discussed above, our sampling and analysis confirmed the presence of new and/or under-collected species, which we detail here. Thanks to our dense sampling in the genus Uvariopsis, we were also able to test species limits, and concluded that several names are actually synonyms, which we detail under each concerned species in the Taxonomic treatment section below.
Morphologically, Uvariopsis sessiliflora stands out from the other Uvariopsis species by having sessile flowers and leaves less than 20 cm long (Up. citrata also has sessile flowers but leaves more than 30 cm long). The only known specimen representing this species is its type specimen (Mildbraed 5239), collected in 1911 in Lomié, a relatively well collected locality of Cameroon (
The recently described species Uvariopsis etugeana Dagallier & Couvreur (
Uvariopsis noldeae Exell & Mendonça is a species endemic to Angola (
Uvariopsis congensis differs from the other Uvariopsis species by its relatively small to medium sized leaves (less than 180 mm long), globose flower buds (vs. conical), and short flowering pedicels (less than 12 mm long). It is a widespread species distributed from Central Africa (Cameroon, Gabon) to East Africa (Uganda, Kenya) (
Finally, the specimens of Uvariopsis congensis from East Africa (
Collection ATBP 666 (African Tropical Biodiversity Programme, MO04937010) collected from the Budongo Forest Reserve in Uganda (as “Uvariopsis sp. nov. Uganda” in Fig.
Uvariodendron
Morphologically, the genus Uvariodendron R.E.Fr. represents the typical Annonaceae flower (
East African Uvariodendron’s: an endless source of botanical discoveries
East Africa is a region that harbours a great plant diversity, with several species being endemic or narrow endemics (
Recently, we described three new Uvariodendron species from the East African coastal forests based on their morphology: Uvariodendron dzomboense Dagallier, W.R.Q. Luke & Couvreur, Uvariodendron mbagoi Dagallier & Couvreur, and Uvariodendron schmidtii W.R.Q. Luke, Dagallier & Couvreur (
In addition to these recently described species, our results reveal another undescribed species. It is represented by four specimens sampled here that were previously identified as Uvariodendron kirkii. These specimens all come from a restricted area in the Kimboza Forest Reserve (Morogoro, Tanzania), and this species is thus described as Uvariodendron kimbozaense Dagallier & Couvreur. Phylogenetically, it is sister to Ud. kirkii although clearly divergent at the molecular level (Fig.

Morphological characters differentiating Uvariodendron kimbozaense Dagallier (A, C, E, G) and Uvariodendron kirkii Verdc. (B, D, F, H) A flowering pedicel with bracts and sepals, side view, note the imbricate sepals B flowering pedicel with bract scar and sepals C flower, with one outer petal and two inner petals gnawed, top view, note the slight transversal curvature of the inner petals D flower, top view, note the ‘boat-shaped’ inner petals E, F simplified representations of the flowers, from above, yellow marking highlights transversal curvature of the petals G, H Simplified representations of the transversal cut of the leaf, by the midrib. A, C Dagallier 49 (type) B, D Dagallier 23. Photos and drawings Léo-Paul Dagallier. Abbreviations: B, bract; brs bract scar; ca, carpel; ip, inner petal; lam, lamina; mlo, lower (abaxial) side of the midrib; mup, upper (adaxial) side of the midrib; op, outer petal; sep, sepal.
Morphological comparison between Uvariodendron kimbozaense and Uvariodendron kirkii (see also Fig.
| Lamina base | Midrib relief above | Flower bud | Bracts | Sepals | Petals size | Inner petals shape | |
|---|---|---|---|---|---|---|---|
| Uvariodendron kirkii | acute to decurrent | slightly impressed (Fig. |
globose, 1.3–6 mm diameter | 1 at base of the pedicel and sometimes 1 on the upper half of the pedicel (Fig. |
3–6.5 mm long, 4–8 mm wide, very broadly to broadly ovate, connivent (Fig. |
10–20 mm long, 6–13 mm wide |
strongly transversally curved (“boat-shape”) (Fig. |
| Uvariodendron kimbozaense | acute to rounded | slightly raised with a central groove all along (Fig. |
globose to oblate, 7–16 mm diameter | 1 at base of the pedicel and from 1 to 4 along the pedicel (Fig. |
6–12 mm long, 12–21 mm wide, depressed ovate, imbricate (Fig. |
16–39 mm long, 9–19 mm wide |
slightly transversally curved (Fig. |
In the Flora Zambesiaca (
Morphological comparison between Uvariodendron dzomboense and Uvariodendron mossambicense. In bold: character unique to the species.
| Lamina shape | Lamina size | Flower bud | Carpels number | |
|---|---|---|---|---|
| Uvariodendron dzomboense | elliptic to narrowly elliptic, acute to slightly decurrent at base, attenuate at apex | 65–132 mm long, 20–45 mm wide | globose, ca. 4.5 mm in diameter, pubescent | 50 to 75 |
| Uvariodendron mossambicense | elliptic, acute to slightly decurrent at base, attenuate at apex | 80–135 mm long, 30–50 mm wide | globose, ca. 6.5 mm in diameter, pubescent | ca. 5 |
East Africa has been relatively well explored botanically, as shown by the completion of the Flora of Tropical East Africa (
The morphological continuum of Uvariodendron fuscum
In the Flora of Cameroon (
Morphologically, Ud. fuscum (Benth.) R.E.Fr., Ud. giganteum (Engl.) R.E.Fr. and Ud. magnificum Verdc. share morphological similarities and differences. They mainly diverge along a morphological continuum by the size of their leaves (lamina length) and the size of their flowers (sepals and petals dimensions, number of carpels), with Ud. magnificum having larger leaves and flowers than Ud. giganteum, with the latter having larger leaves and flowers than Ud. fuscum (Fig.

Morphological continuum in Uvariodendron fuscum: traits comparison for var. fuscum (fusc.), var. giganteum (gig.) and var. magnificum (mag.). Each point represents a measurement. Vertical bars span the minimum and maximum values of the focal traits. Photos, left: leaf of Couvreur 1046; top right: flower, below view, of Couvreur 1046; below right: flower, top view, of Couvreur 990 (CC BY-NC 4.0 Thomas Couvreur).
Uvariodendron fuscum: morphological comparison between var. fuscum, var. giganteum and var. magnificum. In bold: character useful to differentiate the varieties.
| Ud. fuscum | Young branches indumentum | Lamina size | Sepals size | Petals size | Carpels number |
|---|---|---|---|---|---|
| var. fuscum | sparsely pubescent to glabrous | 160–450 mm long, 43–118 mm wide | 11–23 mm long, 13–26 mm wide | 20–42 mm long, 15–26 mm wide | 20 to 70 |
| var. giganteum | with long soft hairs producing a whitish appearance quickly falling off | 357–676 mm long, 84–225 mm wide | 20–30 mm long, 16–26 mm wide | 21–40 mm long, 17–30 mm wide | 50 to 100 |
| var. magnificum | with long soft hairs producing a whitish appearance quickly falling off | 210–750 mm long, 57–215 (250) mm wide | 30–55 mm long, 30–43 mm wide | 50–70 mm long, 36–47 mm wide | 150 to 160 |
Phylogenetically, the specimens of the three species clustered together, with strong support at the crown node and a weak to moderate support at the internal nodes (Fig.
The name Ud. giganteum was already reduced into synonymy with Ud. fuscum in the Flora of Cameroon (
The complexity of Uvariodendron molundense
The species Uvariodendron molundense (Engler & Diels) R.E.Fr. is distributed in Cameroon and Gabon. In 1969, Le Thomas synonymized the names Uvariodendron mayumbense and Uvariodendron letestui with Ud. molundense. Indeed, the position of the flowers along the trunk and branches, previously used to discriminate these species (
Taxonomic revision of Dennettia, Uvariodendron and Uvariopsis
Taxonomic history
Uvariodendron
The names referring to species later placed within Uvariodendron were first described by
In 1930,
Note about the genus name Uva: According to Kuntze, who undertook nomenclatural work between the end of the 19th and the beginning of the 20th centuries, the generic name Uva should be used instead of Uvaria (
Dennetia and Uvariopsis
The genus Uvariopsis Engler was erected in 1899 with the description of Uvariopsis zenkeri Engler (
In parallel, two other genera, morphologically closed to Uvariopsis and Tetrastemma were described: Thonnera De Wild. and Denettia Baker f. The genus Thonnera included a single species Thonnera congolana De Wild. differing from Tetrastemma by flowers having three petals (
In 1933, Robyns & Ghesquière described Uvariopsis vanderystii Robyns & Ghesq., and suggested it was a kind of intermediate species showing characters of Tetrastemma, e.g. cauliflory, and of Uvariopsis, e.g. fused petals (
Morphology and informative characters
Vegetative characters
The genera Dennettia, Uvariodendron and Uvariopsis are trees or shrubs with plagiotropic branches on an orthotropic axis, with indumentum of simple hairs and with no latex or exudate. A distichous phyllotaxis of the primary axis (the trunk) was reported for Ud. pycnophyllum (
The branches can be sparsely pubescent when young, but the hairs generally fall off with age. The dense brown tomentum on the young branches persisting on older branches distinguishes Ud. calophyllum from the other species of Uvariodendron. The species Ud. fuscum (var. giganteum and var. magnificum) also have a characteristic indumentum of long soft hairs producing a whitish appearance on the younger branches, but that quickly disappear in older branches.
In the three genera, the leaves are distichous, simple, entire, exstipulate, with the midrib impressed above, raised below, the secondary veins weakly brochidodromous to brochidodromous and the tertiary veins reticulate. Dennettia has relatively small (less than 160 mm long) elliptic leaves, while Uvariodendron and Uvariopsis have a range of leaf shapes, from elliptic to obovate to oblong, ranging from 65 to 750 mm long. Uvariodendron fuscum var. giganteum has one of the longest leaves of Annonaceae in Africa, up to 60 cm long (
In Uvariodendron, several species (Ud. dzomboense, Ud. gorgonis, Ud. mossambicense and Ud. schmidtii) show ‘eragrostiform’ leaf buds, that are terminal or axillary buds composed of several (generally 5 to 12) distichous and densely pubescent scales (Figs
The scent of crushed leaves and/or bark can be used as a distinctive character. Uvariodendron citriodorum and Up. citrata emit a strong lemon scent (
Inflorescences and flowers
In contrast to sterile parts, Dennettia, Uvariodendron and Uvariopsis have very different fertile parts. In Annonaceae, the inflorescence is a rhipidium, i.e. determinate (sympodial) inflorescence with a terminal flower and lateral cymose and monochasial partial inflorescences (
In Dennettia, the inflorescences are axillary and borne on leafy twigs or borne on old branches, held by a tiny peduncle. They can be composed of up to six flowers, but one or two-flowered inflorescence are the most common. The flowers of Dennettia are bisexual. They are composed of one whorl of three sepals and one single whorl of three petals, which is characteristic of the genus in the Monodoreae. In Annonaceae, single whorls of three petals can be found in Annickia, Annona, and Dasymaschalon (
In Uvariodendron, the inflorescences are axillary and borne on leafy twigs, borne on old branches or borne on the trunk. The axis of the inflorescences are contracted, rendering their interpretation difficult without a detailed morpho-anatomical study. They appear to be rhipidia composed of one or two (in most species), three (Ud. calophyllum, Ud. gorgonis, Ud. mbagoi) or rarely up to 11 (Ud. kimbozaense) flowers. Generally, in multi-flowered rhipidia, the flower in the terminal position is fully developed while the lower flowers on the axis are buds.
Flowers in Uvariodendron are bisexual with one whorl of three sepals and two whorls of three petals, as usually found in Annonaceae (
Uvariopsis has inflorescences borne on trunk in most of the species, but can be axilary to leaves on young branches (Up. congensis, Up. guineensis, Up. lovettiana, Up. oligocarpa, Up. zenkeri). They are generally composed of one to three flowers. Like in Uvariodendron, the peduncle and partial peduncle are contracted, rendering their interpretation as rhipidium difficult. In particular, Up. dioica, Up. submontana and Up. korupensis have inflorescences in clumps of up to 50 flowers on the trunk.
The flower buds of Uvariopsis have different shapes important to discriminate between species (
The fruits of Uvariopsis are composed of several sessile to subsessile monocarps generally cylindrical in shape. Their texture and pubescence can be informative: either smooth and not ridged (Up. congensis, Up. dioica, Up. guineensis, Up. korpensis, Up. lovettiana, Up. oligocarpa) or wrinkled to verrucose (Up. bakeriana, Up. congolana, Up. pedunculosa, Up. solheidii), and either pubescent to tomentose (Up. pedunculosa, Up. oligocarpa, Up. zenkeri) or sparsely pubescent to glabrous.
Pollination and seed dispersal
The pollination biology of Dennettia, Uvariodendron and Uvariopsis is poorly known for most of the species, except for Ud. connivens, Ud. calophyllum, Up. bakeriana and Up. pedunculosa (
In Uvariopsis, the species Up. bakeriana and Up. pedunculosa have been reported as emitting a strong pungent scent, and to be pollinated by flies (Diptera) (
Similarly to pollination biology, data on seed and fruit dispersal of Dennettia, Uvariodendron and Uvariopsis remain scarce. Fruits of Uvariodendron are part of Western gorillas (Gorilla gorilla)’s diet (
Ethnobotany
Uvariodendron and Uvariopsis species are used for food. The ripe fruits of Ud. angustifolium (Jones 3480) and Ud. fuscum (van Andel 3761) are eaten, as well as the young leaves of Ud. connivens (Cheek 5180) and Ud. fuscum (Cheek 5145) that are boiled and used in soup. The bark of Ud. mbagoi (Dagallier 39) is used as infusion for tea or as spice in meat meals. The leaves of Up. citrata (Letouzey 9017) are used to wrap the fish during the cooking to give it an aromatic taste.
Uvariodendron and Uvariopsis species are also part of the pharmacopoeia of several local communities, such as in Benin and Cameroon where Ud. angustifolium, Ud. connivens, Ud. fuscum, Up. bakeriana, Up. dioica and Up. korupensis are used to cure various conditions (
The fruits of Dennettia tripetala, also known as “pepper fruit”, are famous in West Africa for their spicy taste, especially in Nigeria (Fig.

Dennettia tripetala Baker f A flowering branch B flower, one petal removed, showing receptacle C receptacle with stamens and stigmas D stamen, front view E carpel, front view F carpel, longitudinal section showing ovules G flower, top view H flower, side view I fruits collected for local use in Nigeria A–F material of drawings unknown, author of drawing unknown, modified from
Other usages such as firewood and craft were reported for Ud. angustifolium, Up. bakeriana, Up. dioica, and Up. korupensis (
Taxonomic treatment
Ophrypetaleae
Ophrypetaleae , trib. nov.
Type
Ophrypetalum Diels.
Description
Shrubs to trees; indument of simple hairs. Carpels 1–15. Monocarps 1–10, sessile to shortly stipitate, 5 cm long or more, narrowly cylindrical, length:width ratio more than 5.
Included genera
Ophrypetalum Diels (1 species), Sanrafaelia Verdc. (1 species).
Key to the genera of Monodoreae as recognized in this study
| 1 | Midrib of leaf blade clearly raised above; one single large fruit with seeds unordered | 2 |
| – | Midrib of leaf blade impressed above, or slightly raised with a central groove all along the midrib above; fruits with distinct monocarps with uni- or bi-seriate seeds | 3 |
| 2 | Corolla lobes similar and equal in length, forming a distinct tube at the base, margins generally straight | Isolona |
| – | Corolla lobes clearly differentiated into inner and outer petals (but basally fused); the outer ones longer than inner ones, margins generally undulated or crisped | Monodora |
| 3 | Sepals 2 and at least one of: petals 4, flowers unisexual | Uvariopsis |
| – | Sepals 3 and petals 3 or 6 and flowers bisexual | 4 |
| 4 | Sepals 3, petals 3 | 5 |
| – | Sepals 3, petals 6 | 6 |
| 5 | Calyx forming a short and thick receptacular tube (East Africa) | Lukea |
| – | Calyx forming a ring with 3 lobes generally distinct (West Central Africa) | Dennettia |
| 6 | Petals fused at base | 7 |
| – | Petals free | 8 |
| 7 | Petals plicate, transversely folded in bud | Hexalobus |
| – | Petals not plicate, not folded in bud | Asteranthe |
| 8 | Petals in a single whorl, calyx forming a thick receptacular tube | Monocyclanthus |
| – | Petals in two distinct whorls, calyx not forming a receptacular tube | 9 |
| 9 | Receptacle columnar; anther connective reduced to a tuft of hairs | Mischogyne |
| – | Receptacle convex to flat but not columnar; connective not reduced to a tuft of hairs | 10 |
| 10 | Sepals reduplicate-valvate (ridged) in bud | Uvariastrum |
| – | Sepals not reduplicate-valvate (not ridged) in bud | Uvariodendron |
Dennettia
Dennettia tripetala Baker f., Cat. Pl. Oban 5 (1913).
≡ Uvariopsis tripetala
(Baker f.) G.E.Schatz, Novon 13(4): 447 (2003). Type. Nigeria – Edo State • R.E. Dennett 44 (lectotype: K! (K000040959), designated by
Description
Shrub to tree 1–5 m tall, D.B.H unknown; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 2–5 mm long, 1–1.5 mm wide, glabrous. Leaf lamina 72–155 mm long, 30–68 mm wide, length:width ratio 1.9–3.25, elliptic, papyraceous to coriaceous, base acute to decurrent, apex attenuate, acumen 6–13 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 5–10 pairs, brochidodromous to weakly brochidodromous, slightly raised above, raised below; tertiary veins reticulate. Flowers bisexual. Flower buds globose. Inflorescences borne on branches or axilary, composed of 1 to 6 flowers. Peduncle ca. 0–1 mm long, ca. 1 mm in diameter. Flower pedicel 3.5–10 mm long, 1–2.5 mm in diameter, pubescent to sparsely pubescent. Bracts 1 to 3 at base, upper bract 0.5–2 mm long, 1.5–2 mm wide, broadly ovate, pubescent outside, glabrous inside. Sepals 3, 1–3 mm long, 1.5–4 mm wide, broadly ovate, fused up to more than 50% of their length, forming a persistent ring with the 3 lobes generally clearly distinct, curved downward on fruit pedicel, pubescent outside, glabrous inside, color unknown. Petals 3, 5–14 mm long, 4.5–10 mm wide, length:width ratio 1–1.7, broadly ovate to ovate, connivent in bud, free at anthesis, pubescent to sparsely pubescent outside, glabrate inside, light brown to dark red outside, purplish pink inside. Stamens number unknown, 0.5–1 mm long, 0.3–0.5 mm wide, anthers linear, connective prolongation truncate. Carpels 8–30, 2–4.5 mm long, 0.5–2 mm wide, pubescent, free; stigma 0.5–1 mm long, 0.5–1 mm wide, coiled, pubescent. Fruiting pedicel 5–14 mm long, 1–2.5 mm in diameter, pubescent to glabrous. Monocarps, 1–7, 11–32 mm long, 5–18 mm wide, length:width ratio 1.5–2.3, cylindrical, slightly constricted between the seeds when dried, sparsely pubescent to glabrous, greyish green when unripe to pink-reddish when ripe, subsessile; stipe 0.1–3 mm long, 1–2 mm wide, pubescent. Seeds 4–12 per monocarp, biseriate, 7.5–10 mm long, 2–5 mm wide, ellipsoid.
Distribution
Element of the Upper Guinean Domain and Lower Guinean Domain of the Guineo-Congolian Region: Guinea, Sierra Leone, Ivory Coast, Ghana, Benin, Nigeria, Cameroon.
Habitat and ecology
Lowland mature or secondary rain forests, in west Africa reported from drier forests on rocky outcrops (
Phenology
Flowers collected from February to April. Fruits collected from January to July.
Vernacular names
Cameroon: ‘Bushpèpè’ (Westphal 9932). Ivory Coast: ‘Michiti à petites feuilles’ (Aubréville 145). Nigeria: ‘Mmimmi’ in Igbo, ‘Ata Igebere’ or ‘Igberi’ in Yoruba, ‘Opipi’ in Idoma, ‘Imako’ in Urhobo, ‘Ako’ in Bini and Edo, ‘Nkarika’ in Efik and Ibibio (
Uses
Fruits used as spice and leaves, roots, fruits and seeds used as local medicine.
Notes
This species resembles Uvariopsis congensis, Up. oligocarpa and Up. zenkeri in having elliptic leaves generally less than 16 cm long with decurrent base, but it clearly differs by its bisexual flowers having three sepals and three petals.
Preliminary conservation status
This species is distributed from Guinea to Cameroon. Its EOO is estimated at 705,671 km2 and its AOO at 120 km2. Following IUCN criterion B, it would thus be assigned a status of LC.
Additional specimens examined
Benin – Mono • J.-P. Essou 1443 (BENIN, MO, WAG); Aplahoué, Kpédjihoundéhoué; 7°21'N, 1°44'E; 17 Feb. 1999 – Ouémé • A. Akoègninou 2201 (BENIN, WAG); Kétou, Ewè; 7°28'N, 2°35'E; 01 Feb. 1999 – Zou – K. Küppers 2059 (FR, WAG), Noyau Central forêt class.Lama; 6°57'N, 2°07'E; 28 Feb. 1998. Cameroon – South-West Region • D.W. Thomas 5661 (K, MO, P, WAG), Nganjo. West bank of Meme River on Kumba Mbonge road; 4°33'N, 9°24'E; alt. 50 m; 25 Feb. 1986 • E. Westphal 9932 (WAG), Victoria; 4°01'N, 9°12'E; 04 Apr. 1978 • G.W.J. Mildbraed 10515 (K), Likomba – Pflangzung, 15–35 km NE of Victoria; 4°06'N, 9°20'E; alt. 50 m; 18 Oct. 1928 • T.D. Maitland 626 (K); Fako, Victoria (= Limbe), Bonjongo; 4°06'N, 9°08'E; alt. 600 m; Apr. 1929 • T.D. Maitland 627 (P), Victoria (= Limbe), Bonjongo; 4°06'N, 9°08'E; 1929 • T.D. Maitland s.n (K), Mt Cameroon; 4°15'N, 9°13'E; alt. 920 m; 1930 • T.D. Maitland s.n.14 (K), Mt Cameroon, Buea area; 4°12'N, 9°11'E; alt. 975 m; 1930. Ghana – Ashanti Region • J.K. Morton A3394 (K), Nyinahin range, W of Kunasi; 6°36'N, 2°07'W; alt. 700 m; 07 Jun. 1958 – Central Region • J.B. Hall GC43611 (K, WAG), Ojobi near Awutu; 5°29'N, 0°32'E; 29 Feb. 1972 • J.K. Morton s.n (K), About 1 mile N of Ojubi, Senya Beraku; 5°23'N, 0°29'E; 24 May. 1953 – Unknown major area • J.B. Hall GC43276 (K), Yaugwu F.R; alt. 460 m; 19 Mar. 1972. Guinea – Kindia • D. Molmou 603 (P), Layah, Sousoude. Just avant Tonkoyah; 9°43'39.3"N, 13°12'26.6"W; alt. 12 m; 06 Jul. 2013. Ivory Coast – Bouaflé • C.C.H. Jongkind 4356 (MO, WAG), Parc National de la Marahoue. Near south border; 6°59'N, 6°10'W; alt. 250 m; 11 Feb. 1998 – Bouaké • L. Aké Assi 18031 (G), Forêt de Bamoro; 7°48'N, 5°05'W; 07 Jun. 1989 – Bouna • C. Geerling 2222 (K, WAG), Iringou (Gawi-Bania confl. Bamago); 8°58'N, 3°39'W; 15 Mar. 1968 • P.P. Poilecot 1224 (G), P.N. Comoé Sud; 8°45'N, 3°35'W; 13 Apr. 1986 • P.P. Poilecot 4164 (G, MO), P.N. Comoé Sud; 8°45'N, 3°35'W; 13 Apr. 1986 – Danané • A.J.M. Leeuwenberg 2999 (K, L, MO, P, WAG), 2 km E of Danané; 7°16'N, 8°07'W; alt. 410 m; 06 Mar. 1959 – Oumé • A. Le Thomas 27 (P), Lamto-Station, Fleuve Bandama; 6°13'N, 5°01'30"W; 07 Jul. 1985 • A. Le Thomas 97 (P), Lamto-Station, Bandama; 6°13'N, 5°01'30"W; 26 Jul. 1985 – Sassandra • A. Aubréville 1957/163 (BR, K, P); 4°59'N, 6°08'W; 07 Mar. 1957 – Soubré • L. Aké Assi 9490 (G, P), Foret du Bassin de la Lobo; 6°07'N, 6°48'W; 22 Feb. 1967 – Vavoua • F.N. Kouamé 1470 (CSRS, G), F.C. du Haut-Sassandra, Centre. forêt très dégradée, relevé FNK20; 7°07'N, 7°00'W; 07 Apr. 1995 – Unknown major area • A. Aubréville 145 (K, P); 7°37'26.4"N, 5°33'14.4"W; 28 Feb. 1957 • L. Aké Assi 13255 (G), Mont Niénokoué; 5°23'N, 7°10'W; 19 Jan. 1976 • L. Aké Assi 9543 (G, K, P), Mt Mafa; 5°52'N, 4°04'W; 14 Mar. 1967. Nigeria – Anambra State • J.C. Okafor FHI35869 (K), Onitsha; Nnwei; Ukpor – Nzagha village; 6°50'N, 6°50'E; 22 Oct. 1956 – Cross River State • C.F.A. Onochie FHI36331 (K), Calabar, Ikot Efanga, c. 12 miles from Calabar on the Oban road; 4°58'N, 8°29'E; 09 Feb. 1957 – Oyo State • C.E. Darter 41837 (K), Olokemeji Forest Reservee; agbola pool, Ogun River; 7°25'N, 3°32'E; 06 Apr. 1958 – Unknown major area • J.O. Ariwaodo ARS1184 (K), Munta Village. Sierra Leone – Western Area • J.K. Morton SL2013 (FHI, IFAN, K, SL, WAG); Western Area, near York by Whale Bay, Peninsula; 8°17'N, 13°11'W; 02 May. 1965.

Uvariodendron
= Uvaria section Uvariodendron, Engl. & Diels in Engl. Afr. Pfl. VI 8 (1901).
Type species
Uvariodendron fuscum (Benth.) R.E.Fr. (≡ Uvaria gigantea Engl.).
Description
Shrub to trees 2–30 m tall, D.B.H. 1–35 cm; young branches pubescent to tomentose to glabrous, old branches slightly tomentose to glabrous. Leaves with margin flat to slightly revolute. Petiole 2.5–35 mm long, 1–9 mm wide, pubescent to tomentose to glabrous. Leaf lamina 65–750 mm long, 20–250 mm wide, length:width ratio 2.1–6.2, elliptic to obovate to oblong, coriaceous, base acute to decurrent to rounded to subcordate, apex acute to acuminate, acumen 0.5–38 mm long; surface above glabrous, surface below pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, pubescent to glabrous below; secondary veins 8–41 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk, on branches or axillary, composed of 1–11 sessile to pedicellate flowers. Flower pedicel 0–65 mm long, 1–9 mm in diameter, pubescent to glabrous. Flowers bisexual, buds globose to ovoid, sessile to pedicellate, 2–22 mm high, 4–30 mm in diameter, pubescent. Bracts 1 to 6, upper bract 1–35 mm long, 1–50 mm wide, depressed ovate to ovate, generally adpressed, enclosing the bud or semi-clasping the pedicel, pubescent outside, glabrous to pubescent inside. Sepals 3, 3–55 mm long, 2.5–43 mm wide, valvate or imbricate, free or fused at base, pubescent outside, glabrous inside, green to brown. Outer petals 3, 10–70 mm long, 9–47 mm wide, length:width ratio 0.9–2.5, ovate to elliptic, pubescent outside, puberulent to glabrous inside, cream to brown to wine red outside, cream with dark red at base to wine red inside. Inner petals 3, 10–53 mm long, 5–38 mm wide, length:width ratio 0.9–2.4, elliptic to obovate, pubescent outside, puberulent to glabrous inside, cream to brown to wine red outside, cream with dark red at base to wine red inside. Stamens 200 to 3000, 1–5 mm long, 0.1–0.7 mm wide, anthers linear, connective prolongation truncate. Carpels 5 to 160, 1–7 mm long, 0.5–2.2 mm wide, pubescent, free; stigma 0–2 mm long, 0.5–2 mm wide, coiled, pubescent, generally covered with an exsudate at anthesis. Fruiting pedicel 0–31 mm long, 2–8 mm in diameter, pubescent. Monocarps 1 to 80, 17–90 mm long, 4.5–32 mm wide, length:width ratio 1.3–11, generally cylindrical, pubescent to glabrous, green to orange to red to dark-blue black; sessile to stipitate; stipe 0–12 mm long, 1–10 mm wide, pubescent to glabrous. Seeds 1–18 per monocarp, uniseriate to biseriate, 3 to 29 mm long, 2–11 mm wide, ellipsoid to semicircular.
Key to Uvariodendron species
| 1 | Largest leaves with lamina ≤ 350 mm long | 2 |
| – | Largest leaves with lamina > 350 mm long | 13 |
| 2 | Largest leaves with lamina < 160 mm long | 3 |
| – | Largest leaves with lamina > 160 mm long | 5 |
| 3 | Plant with strong bergamot scent (bark and crushed leaves), leaves stiff, between coriaceous and cartilaginous, apex acute to shortly acuminate; monocarps cylindrical and tomentose with regular tufts of high hair density (Tanzania) | Ud. mbagoi |
| – | Plant without bergamot scent, leaves coriaceous, apex attenuate; monocarps cylindrical and glabrous or ovoid and densely pubescent | 4 |
| 4 | Flower buds ca. 4 mm in diameter, covered by ca. 6 velutinous bracts 5–6 mm long; flowers with ca. 50–75 carpels; monocarps ovoid, densely pubescent, sessile (Kenya) | Ud. dzomboense |
| – | Flower buds ca. 6 mm in diameter, covered by 2–5 velutinous bracts ca. 4 mm long; flowers with ca. 5 carpels; monocarps cylindrical, glabrous, with stipe ca. 12 mm long (Mozambique) | Ud. mossambicense |
| 5 | Sepals fused at base over > 10% of their length | 6 |
| – | Sepals free or fused at base over ≤ 10% of their length, connivent or imbricate | 9 |
| 6 | Plant with a strong lemon scent; leaves narrowly elliptic, base acute to cuneate, flower sessile with petals 15–25 mm long and 5–15 mm wide (West and Central Africa) | Ud. angustifolium |
| – | Plants with no lemon scent; leaves elliptic to oblong to obovate, base acute to decurrent; flower pedicellate, or flower sessile with petals 20–36 mm long and 15–19 mm wide | 7 |
| 7 | Plant with bark reddish peeling off; flower bud globose to ovoid, sessile, > 6 mm high; flower with sepals ≥ 10 mm long and wide (Tanzania) | Ud. pycnophyllum |
| – | Plant with bark not reddish and not peeling off; flower bud globose, subsessile to pedicellate, ≤ 6 mm high; flower with sepals < 8 mm long | 8 |
| 8 | Leaves 170–350 mm long; flowers with sepals 3–5 mm long, outer petals elliptic 15–25 mm long, carpels 20 to 40; fruits with 6 to 11 stipitate (stipe 5–12 mm) monocarps (West Africa) | Ud. occidentale |
| – | Leaves 159–188 mm long, flowers with sepals 5.5–7 mm long, outer petals broadly obovate 11–12 mm long, carpels ca. 7; fruits with 3 to 5 sessile monocarps (Kenya) | Ud. schmidtii |
| 9 | Leaves obovate, base decurrent, apex acuminate; flowers with bract ca. 10 mm long, ca. 30 carpels (Gabon) | Ud. pilosicarpum |
| – | Leaves elliptic to oblong to obovate, base acute to decurrent or acute to rounded, apex acute to attenuate; flowers with bract ≤ 6 mm long, < 20 carpels | 10 |
| 10 | Plant with a strong aniseed smell; flowering pedicels 15–65 mm (Kenya, Tanzania) | Ud. anisatum |
| – | Plant with no aniseed smell; flowering pedicels ≤ 28 mm | 11 |
| 11 | Fresh leaves with strong lemon scent when crushed, leaves ≥ 250 mm (Central Africa) | Ud. citriodorum |
| – | Leaves with no lemon scent when crushed, leaves < 220 mm | 12 |
| 12 | Leaves 140–220 mm long with a midrib slightly raised above with a central groove all along; flower with imbricate sepals 6–12 mm long, and elliptic petals 16–39 mm long with a slight transversal curvature (Tanzania) | Ud. kimbozaense |
| – | Leaves 70–190 mm long with a midrib slightly impressed above; flower with connivent sepals 3–6.5 mm long, ovate petals 10–20 mm long with a strong transversal curvature (Kenya, Tanzania) | Ud. kirkii |
| 13 | Young branches, petioles, and midrib below the lamina covered with a brown tomentum, generally persisting on older branches (West and Central Africa) | Ud. calophyllum |
| – | Young branches, petioles, and midrib below the lamina pubescent to glabrous or covered with long soft hairs quickly falling off | 14 |
| 14 | Leaves with rounded to subcordate base (Tanzania) | Ud. usambarense |
| – | Leaves with acute to rounded base | 15 |
| 15 | Monocarps with l:w ratio ≥ 5, very narrowly cylindrical, torulose to torose (i.e. very strongly constricted between the seeds); seeds < 10 mm long (East Africa) | Ud. gorgonis |
| – | Monocarps with l:w ratio < 3.5, cylindrical, not constricted between the seeds; seeds > 10 mm long | 16 |
| 16 | Young branches glabrous; flowering pedicel ≥ 10 mm, petals wine red outside and inside; monocarps sparsely pubescent to glabrous (Central Africa) | Ud. connivens |
| – | Young branches pubescent to glabrous; flower pedicel ≤ 15 mm, petals wine red cream to light yellow outside, cream with dark red steak inside; monocarps pubescent to sparsely pubescent | 17 |
| 17 | Bracts 3–8 mm long and 3–10 mm wide; sepals free, sometimes fused at base, generally imbricate, 5–9 mm long and 5–10 mm wide (Central Africa) | Ud. molundense |
| – | Bracts 8–35 mm long and 10–50 mm wide; sepals fused at base over 20–50% of their length, 11–55 mm long and 13–43 mm wide | Ud. fuscum (18) |
| 18 | Young branches sparsely pubescent to glabrous; sepals 11–23 mm long and 13–26 mm wide; carpels 20 to 70; fruiting pedicel ca. 5 mm long (Central Africa) | Ud. fuscum var. fuscum |
| – | Young branches with long soft hairs producing a whitish appearance quickly falling off; sepals 20–55 mm long and 16–43 mm wide; carpels 50 to 160; fruiting pedicel ≥ 9 mm long | 19 |
| 19 | Flowering pedicels 0–7.5 mm long, sepals 20–30 mm long and 16–26 mm wide, petals 21–40 mm long and 17–29 mm wide, carpels 50 to 100 (Central Africa) | Ud. fuscum var. giganteum |
| – | Flowering pedicels 10–15 mm long, sepals 30–55 mm long and 30–43 mm wide, petals 50–70 mm long and 36–47 mm wide, carpels 150 to 160 (Uganda) | Ud. fuscum var. magnificum |
Species descriptions
Uvariodendron angustifolium
≡ Uvaria angustifolia Engl. & Diels, Notizbl. Königl. Bot. Gart. Berlin 2: 295 (1899); Uva angustifolia (Engler & Diels) Kuntze, Deutsche Bot. Monatsschr. xxi. 173 (1903). Type. Cameroon – South-West Region • A. Staudt 742a (holotype: B! (B 10 0153115)), Johann Albrechts-Höhe; 4°10'N, 9°12'E; alt. 400 m; 20 Mar. 1896.
Description
Shrub to tree 3–12 m tall, D.B.H. unknown; young branches sparsely pubescent to glabrous, old branches glabrous; plant with strong aromatic smell. Leaves with strong lemon smell when crushed. Petiole 3–7.5 mm long, 1–2 mm wide, pubescent to glabrous. Leaf lamina 100–199 mm long, 30–58 mm wide, length:width ratio 3–4, narrowly elliptic, coriaceous, base acute to cuneate, apex acute to broadly acuminate, acumen 11–14 mm long; surface above glabrous, surface below pubescent to glabrous when young, pubescent at base to glabrous when old; midrib slightly impressed above, raised below, glabrous above, pubescent to glabrous below; secondary veins 8–14 pairs, weakly brochidodromous, slightly impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and branches, composed of 1 (sub)sessile flower. Flower pedicel 0–6 mm long, 2–3 mm in diameter, velutinous. Flowers bisexual, buds globose, sessile to shortly pedicellate, 5–8 mm high, 6–8 mm in diameter, sericeous. Bracts 1 to 6, upper bract 6–11 mm long, 9–15 mm wide, ovate, appressed, enclosing the bud, sericeous outside, glabrous inside. Sepals 3, 9–13 mm long, 9–13 mm wide, fused at base, slightly imbricate at mid-length, sericeous outside, glabrous inside, color unknown. Outer petals 3, 15–25 mm long, 9–15 mm wide, length:width ratio 1.13–1.78, elliptic to oblong, velutinous outside, glabrous inside, yellowish brown outside, crimson with cream margins inside. Inner petals 3, 15–22 mm long, 5–10 mm wide, length:width ratio 1.9–3, obovate, velutinous outside, glabrous inside, yellowish brown outside, crimson with cream margins inside. Stamens 200 to 300, 3–3.5 mm long, 0.1–0.5 mm wide, anthers linear, connective prolongation truncate. Carpels 7 to 30, 3–4.5 mm long, 1–1.5 mm wide, pubescent, free; stigma 1–2 mm long, 0.8–1.5 mm wide, coiled, pubescent, covered with an exudate at anthesis. Fruiting pedicel ca. 5 mm long, 3.5 mm in diameter, pubescent. Monocarps 2 to 10, 23–40 mm long, 17–30 mm wide, length:width ratio ca 1.3, ellipsoid, smooth, sparsely pubescent to glabrous, yellow, with strong lemon smell; shortly stipitate, stipe 1–3.5 mm long, 1–2.5 mm wide, pubescent. Seeds 9 to 18 per monocarp, biseriate, 21 to 28 mm long, ca. 10 mm wide, 2–4 mm thick, semicircular.

Uvariodendron connivens (Benth.) R.E.Fr. A leaf B flower, two outer and one inner petal removed C outer petal inner view D stamen, front view. Uvariodendron angustifolium (Engl. & Diels) R.E.Fr. E leaves F flower, semi top view G flower, bottom view showing bracts H flower, bottom view, bracts removed I outer petal, outer view J stamen, front view K carpel, front view and detail of ovules. A–D from Mann 1159 A–K from Vigne 1610. Drawings by Hélène Lamourdedieu, Publications Scientifiques du Muséum national d’Histoire naturelle, Paris.
Distribution
Endemic to Upper Guinean Domain and Lower Guinean Domain of the Guineo-Congolian Region: Cameroon, Ghana, Ivory Coast, Nigeria. This species has also been reported in Benin (
Habitat and ecology
Lowland mature or secondary rain forests. Altitude: 152–400 m a.s.l.
Phenology
Flowers collected from February to March and November to December. Fruits collected from January to May.
Vernacular names
Ghana: ‘Esuno Kodu’ or ‘Bomborgo Kodu’ in Ashanti (Vigne 1610).
Uses
The fruits are used to flavour the soup (Jones FHI 3480).
Notes
This species can be easily distinguished from the other Uvariodendron species by its narrowly elliptic leaves. It also emits a strong citrus scent when fresh, a characteristic shared with other Annonaceae species: Uvariodendron citriodorum (Le Thomas) Dagallier & Couvreur and Uvariopsis citrata Couvreur & Niangadouma.
Preliminary conservation status
This species is distributed from Cameroon to Ivory Coast. Its EOO is estimated at 186,201 km2 and its AOO at 52 km2. It occurs in ca. 10 locations, but the last collection dates back more than 20 years and two thirds of the collections are older than 50 years, suggesting a decline in the number of locations and in AOO. We thus assign a preliminary conservation status of Vulnerable VU under criterion B2ab(ii,iv).

Additional specimens examined
Cameroon – Central Region • R.W.J. Keay FHI37524 (FHI, K), between Bafia and Likoko; 4°21'N, 9°18'E; 05 Feb. 1958 – South-West Region • D.W. Thomas 6087 (MO, YA); Meme, Vicinity of Lake Barombi, Kumba; 4°39'N, 9°24'E; alt. 300 m; Apr. 1986 • D.W. Thomas 7018 (YA), Cocoa farms along the road between Konye and Bakole; 4°39'N, 9°25'E; alt. 300 m; 25 May. 1987 – Unknown major area • A. Staudt 642 (BM, G, K), Kumba; 4°38'N, 9°25'E; 1896. Ghana – Ashanti Region • C. Vigne 1610 (K, P), Pra River, ashanti; 6°33'54.36"N, 0°57'21.6"E; alt. 152 m; Feb. 1929 – Western Region • N.K. Lovi 3964 (K, P), New Tafo E.P; 6°13'N, 0°22'E; 03 Dec. 1954. Ivory Coast – Bouaflé • C.C.H. Jongkind 4368 (IAGB, MO, WAG), Parc National de la Marahoue; 7°02'N, 6°06'W; alt. 270 m; 13 Feb. 1998 – Daoukro • A.J.B. Chevalier s.n (P), Morénou: environs du poste de Bangouanou; 6°39'N, 4°12'W; 26 Nov. 1909. Nigeria – Edo State • B.O. Daramola 94/337 (K, MO), Iyekoriomwon, Edo State, about 35 kilometers from Uyo (Ugo ?); 6°05'19.05"N, 6°00'04.46"E; 09 Jan. 1994 – Ondo State • A.P.D. Jones FHI3480 (K); Owo District, Owo forest reserve about 4 miles North of Igbatoro crossing; 5°33'36.36"N, 6°56'06"E; 19 Apr. 1943 • F. Anakwense FHI19702 (K); Akure, Akure Forest Reserve; 7°19'N, 5°02'E; 11 Dec. 1950 – Oyo State • D. Gledhill 817 (K); Ibadan District, Gambari Forest Reserve; 7°08'N, 3°50'E; 02 Feb. 1968.
Uvariodendron anisatum
Type
Kenya – Nairobi • B. Verdcourt 526 (holotype: EA! (EA000002461); isotypes: K! (K000198890, K000198891), PRE); Nairobi District, in Karura Forest and Thika Gorge; 1°14'S, 36°49'E; 24 Jun. 1951.
Description
Shrub to tree 3.5–15 (22) m tall, D.B.H. up to 10 cm; young branches sparsely pubescent to glabrous, old branches glabrous; plant with aniseed smell. Leaves reddish pink when young, dark green when old. Petiole 3–9 mm long, 1.5–4 mm wide, sparsely pubescent to glabrous. Leaf lamina 80–313 mm long, 45–187 mm wide, length:width ratio 2.3–3.1, elliptic to obovate, coriaceous, base acute to decurrent, apex acute to attenuate, surface above glabrous, surface below sparsely pubescent to glabrate when young, glabrous when old; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 12–21 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences clustered on the trunk or borne on branches, composed of 1–3 flowers. Flower pedicel 15–65 mm long, 1–2 mm in diameter, pubescent. Flowers bisexual, buds globose, pedicellate, 3.5–6 mm high, 4.5–6 mm in diameter, sericeous. Bracts 1 to 2 at base and 1 towards the upper half of the pedicel, upper bract 2–4.5 mm long, 3–8 mm wide, clasping the pedicel, velutinous outside, glabrous inside. Sepals 3, 3.5–6 mm long, 4–8 mm wide, valvate to fused at base over 10 % of their length, velutinous outside, glabrous inside, color unknown. Outer petals 3, 10–23 mm long, 9–16 mm wide, length:width ratio 1–1.6, broadly ovate to ovate, velutinous outside, glabrous inside, cream outside, cream inside. Inner petals 3, 12.5–15 mm long, 5–9 mm wide, length:width ratio 1.5–2, obovate, shortly velutinous outside, glabrous inside, cream outside, cream inside. Stamens 400 to 500, 1.5–2.5 mm long, 0.1–0.5 mm wide, anthers linear, connective prolongation truncate. Carpels 7 to 12, 2–5 mm long, 1–1.5 mm wide, pubescent, free; stigma 1–2 mm long, 1–1.5 mm wide, coiled, pubescent. Fruiting pedicel 21–59 mm long, 2–3 mm in diameter, sparsely pubescent to glabrous. Monocarps 3 to 14, 38–70 mm long, 13–20 mm wide, length:width ratio 2.3–3.5, cylindrical, longitudinally ridged, slightly constricted between the seeds, slightly acuminate, sparsely pubescent, dark olive green to dark-blue black; sessile to shortly stipitate, stipe 0–2 mm long, 3–4.5 mm wide, sparsely pubescent. Seeds 8–18 per monocarp, biseriate, 10 to 12 mm long, 5–8 mm wide, ellipsoid to semicircular.

Uvariodendron anisatum Verdc A entire specimen sheet with young branch and leaves B fruit with four monocarps (one detached) C stamen, side view D flower and pedicel attached to bark clump, semi-bottom view E detail of flower, bottom view F detail of flower showing sepals, one outer petal removed, semi-top view. A Verdcourt 406 B Verdcourt 674 D–F Faden 74/886. Photos Léo-Paul Dagallier.
Distribution
Endemic to Somalia-Masai Region: Kenya and Tanzania.
Habitat and ecology
Submontane rain forests (sometimes dry forests), often along streams. Altitude: 300–1080 m a.s.l.
Phenology
Flowers collected in January, March, from June to July and from November to December. Fruits collected in January, March, from May to July and from November to December.

Vernacular names
Kenya: ‘Mutongu’ in Meru (Parnell 13340).
Notes
This species is characterized by a strong aniseed smell (lightly to not noticeable on herbarium specimens), a character unique in the Annonaceae family to my knowledge. Apart from the smell, it closely resembles Uvariodendron kirkii and can hardly be differentiated on dry sterile material. In flowers, it can be differentiated from Ud. kirkii by its flower pedicel generally longer (15–65 mm, vs. 5–28 mm), its relatively flat petals with a slight transversal curvature (vs. “boat-shaped” petals, i.e. with a strong transversal curvature). Note that the petals curvature is not easy to rely on dry specimen. In fruits, it can be differentiated with a greater fruit pedicel than Ud. kirkii (21–59 mm long, vs. 7–22 mm long). Verdcourt also used the monocarps to differentiate the 2 species, the monocarps of Ud. anisatum being dark blue and with a slight longitudinal ridge, and the monocarps of Ud. kirkii being dull-orange and without any pseudosuture (
Conservation status
This species has been assessed as Vulnerable VU under criteria B2ab(ii,iii,iv,v) (
Additional specimens examined
Kenya – Central • E. Battiscombe 1285 (EA, K); Kiambu District, Kiambu Forest; 1°10'S, 36°49'E; alt. 1700 m; 17 Jun. 1924 • J.L. Moon 811 (K); Kiambu District, Kyambu; 1°11'48.48"S, 36°51'32.76"E; alt. 1737 m; 29 Jul. 1913 – Eastern • H.J. Beentje 4089 (EA); Meru, Kijegge Hill; 0°17'N, 37°57'E; alt. 1400 m; 08 Mar. 1989 • I. Malombe 1368 (EA), Ngaia Forest, Kiegoi village, NE border; 0°14'N, 37°55'E; alt. 1170 m; 22 May. 2008 • L.E. Parnell EA13340 (K); Meru, Lower Imenti Foest Reserve; 0°07'14.88"N, 37°41'52.08"E; 24 Aug. 1965 • M.A. Brunt 1546 (K), Embu Meru road; 0°15'43.26"N, 37°38'23.43"E; 16 Feb. 1964 • R.B. Faden 70/120 (EA, K); Meru District, Lower Imenti Forest, 6 mils. from Meru along Meru – Mikinduri Road; 0°06'N, 37°45'E; alt. 1190 m; 01 Mar. 1970 • S.A.L. Smith 287 (K); Embu District, Kiangombe, northern slopes; 0°33'52.56"N, 37°42'27.72"E; alt. 1550 m; 30 Nov. 2000 • W.R.Q. Luke 7123 (EA); Meru, Ngaia Forest, camp 142; 0°19'N, 38°02'E; alt. 1080 m; 22 Nov. 2000 – K4 Central • H.J. Beentje 1830 (EA, WAG), Nairobi, E. Karura forest near Kiambu R; 1°14'30"S, 36°49'30"E; alt. 1600 m; 13 Jan. 1985 • R.B. Faden 74/886 (K, WAG), Meru District. Lower Imenti Forest, on Meru-Mikinduri Road; 0°04'N, 37°45'E; alt. 1190 m; 26 Jun. 1974 – Nairobi • B. Verdcourt 3689 (K); Nairobi District, Karura Forest, by Ruaraka R. to W. of main Thika highway; 1°14'S, 36°49'E; 21 Jul. 1963 • B. Verdcourt 406 (B, K, K); Nairobi District, Thika Road House; 1°15'59.57"S, 36°50'04.08"E; alt. 1676 m; 24 Dec. 1950 • B. Verdcourt 674 (K); Nairobi District, Karura Forest; 1°14'S, 36°49'E; 29 Jun. 1952 • E. Polhill 324 (K); Nairobi District, Karura Forest; 1°14'S, 36°49'E; 10 Jan. 1961 • V.G.L. van Someren 95 (G, K); Nairobi District, Karura Forest; 1°14'S, 36°49'E; 13 Mar. 1940. Tanzania – Kilimanjaro • K.B. Vollesen 96/15 (K), Mkomazi Game Reserve, Ibaya Hill; 3°59'S, 37°47'E; alt. 1350 m; 03 Jun. 1996 • R.A. Abdallah 96/86 (K), Mkomazi National Park, Mkomazi Game Reserve, Maji Kununua Ridge; 3°53'S, 37°48'E; alt. 1500 m; 07 Jun. 1996 – Pwani • R.P. Sacleux 212 (P), Mandéra (Zanguebar); 6°12'51.22"S, 38°22'37.73"E; Jan. 1889 – Tanga • R.C. Wingfield 2880 (K), Msangasi Forest Reserve, 50 km S of Korogwe; 5°36'55.14"S, 38°27'06.26"E; alt. 300 m; 31 Mar. 1975.
Uvariodendron calophyllum
= Uvaria gigantea Engl., Notizbl. Königl. Bot. Gart. Berlin 2: 292. (1899) (quoad specimen Zenker 1738).
Type
Cameroon – South Region • G.A. Zenker 2344 (holotype: B! (B100153116); isotypes: BM! (BM000636669), E!(E00704856), G! (G00412241), GOET! (GOET005732), HBG! (HBG502513), K! (K000198796, K000198797), M! (M0107940), P! (P00362658, P00362659, P00362661), S! (S07-13393, S07-13396), WAG! (WAG.1418666)), Bipindi; 3°05'N, 10°25'E; 1901.
Description
Tree 2–20 m tall, D.B.H. 1–32.1 cm; young branches tomentose, old branches slightly tomentose to glabrous. Leaves reddish when young, dark green when old. Petiole 4–25 mm long, 2–9 mm wide, tomentose. Leaf lamina 258–765 mm long, 61–248 mm wide, length:width ratio 2.2–4.9, obovate, coriaceous, base rounded to slightly truncate, apex acuminate, acumen 6–32 mm long; surface above glabrous, surface below pubescent to glabrous; midrib impressed above, raised below, glabrous above, tomentose below; secondary veins 20–41 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and branches, composed of 2–3 (sub)sessile flowers. Flower pedicel 0–11 mm long, 4–9 mm in diameter, tomentose. Flowers bisexual, buds globose, sessile, 7–17 mm high, 7–30 mm in diameter, tomentose. Bracts 1 to 6, upper bract 10–23 mm long, 10–40 mm wide, ovate, appressed, enclosing the bud, tomentose outside, glabrous inside. Sepals 3, 10–27 mm long, 10–26 mm wide, imbricate to fused at base, tomentulose outside, glabrous inside, dark brown outside, reddish inside. Outer petals 3, 15–46 mm long, 10–33 mm wide, length:width ratio 0.9–1.5, broadly elliptic to elliptic, tomentulose outside, glabrous inside, cream outside, cream with dark red at base inside. Inner petals 3, 14–41 mm long, 11–32 mm wide, length:width ratio 1.2–1.9, obovate, shortly tomentulose outside, glabrous inside, cream outside, dark red with cream margins inside. Stamens around 3000, 3.5–4.9 mm long, 0.1–0.5 mm wide, anthers linear, connective prolongation truncate. Carpels 35 to ca. 150, 2–5 mm long, 0.5–2 mm wide, pubescent, free; stigma 1–2 mm long, 0.5–2 mm wide, coiled, velutinous, covered with an exudate at anthesis. Fruiting pedicel 1–7 mm long, 4–7 mm in diameter, densely pubescent. Monocarps 3 to 35, 27–55 mm long, 9–25 mm wide, length:width ratio 1.8–3.7, ellipsoid to obovoid, tomentose, brown; sessile to shortly stipitate, stipe 0–10 mm long, 1.5–3 mm wide, tomentose. Seeds 7–13 per monocarp, biseriate, ca. 13 mm long, 8–9 mm wide, semicircular.

Uvariodendron calophyllum R.E.Fr. A leaf apex upper side B detail of young branch and leaf base, upper side, note the brown tomentum C detail of young branch and leaf base, lower side D leaf E flower fully open, top view F flower with inner petals still attached to each other, semi-top view G fruit with many monocarps (dried) H flower buds, note the brown tomentum. A, D, H Couvreur 1157 B, C Couvreur 999 E, F Couvreur 1013 G Letouzey 14020. Photos A–F, H Thomas Couvreur, G Léo-Paul Dagallier.
Distribution
Element of the Upper Guinean Domain and Lower Guinean Domain of the Guineo-Congolian Region: Cameroon, Gabon, Ghana, Ivory Coast, Nigeria.
Habitat and ecology
Lowland mature or old secondary rain forests, near streams. Altitude: 30–140 m a.s.l.
Phenology
Flowers collected from December to May. Fruits collected from March to May, from July to August, and in November.
Vernacular names
Cameroon: ‘Ebom Bulu’ (Parren 223) or ‘Ebom Afame’ (Parren 68) in Bulu/Ewondo.
Notes
This species is close to Uvariodendron connivens and Ud. fuscum var. giganteum by having large obovate leaves (more than 25 cm long). However, it can easily be distinguished from these three other species as it presents a tomentum of short dense brown matted hairs on the young parts (branches, petioles and lower midrib), producing a brown appearance with whitish reflections. The flower buds and outer petals are also covered outside with a similar brown and dense tomentum. The older parts of the plant are generally glabrous but can also present remnants of the tomentum.

Conservation status
This species is widespread, distributed from Ivory Coast to Gabon. It has been previously assessed as Least Concern LC (
Additional specimens examined
Cameroon – Central Region • J.N. Asonganyi 421 (P, YA), Ndikiniméki, at Sonossi 26 km W. of Ndikinimeki. Map 1/200 000; 4°45'44.24"N, 10°36'29.26"E; 29 Mar. 1982 – Littoral • A.J.M. Leeuwenberg 5282 (BR, K, MO, P, PRE, WAG), 8 km W of Masok; 4°08'N, 10°24'E; alt. 400 m; 31 Mar. 1965 • A.J.M. Leeuwenberg 5282a (K, P, WAG), 8 km W. of Masok; 4°08'N, 10°24'E; alt. 400 m; 31 Mar. 1965 • R.G. Letouzey 12352 (P, YA), colline entre Tcherikoy et Sokelle II(30 km NW Eseka) – feuille IGN 1/200 000 EDEA; 3°48'18.03"N, 10°34'32.35"E; 14 Dec. 1973 – North-West Region • E.U. Ujor FHI29281 (FHI, K); Mentchum, Nkom-Wum, Bamenda Prov., Wum Distr., Nkom • Wum F.R. on German old road leading to Timber Camp by the left; 6°16'N, 10°08'E; 03 Jul. 1951 • R.G. Letouzey 14020 (P, WAG, YA), Piste Baji-Tumbo, entre Baji et rivière Chonogbonbon, 55 km NNE Wum; 6°45'N, 10°10'E; 12 Jul. 1975 – South Region • G.A. Zenker 1738 (B (B101178582), BM (BM000636667), COI (COI00004925), E (E00718583), G (G00412236), HBG (HBG502512), K (no barcode), LE (LE00012448), LECB (LECB0000053), M (M0239941), P (P06901474, P06901466, P06901467), WAG (WAG.1418665), WU (WU0025882, WU0025883), Z (Z-000056111, Z-000056112)), Bipindi; 3°05'N, 10°25'E; 1898 • G.A. Zenker s.n (P), Bipinde, Bipindi; 3°05'N, 10°25'E; Apr. 1903 • M.P.E. Parren 223 (KRIBI, WAG), About 7 km NE of Ebom. Plot 13, subplot 84, tree 6, coordinates: X = 7, Y = 3 m; 3°07'N, 10°45'E; alt. 500 m; Aug. 1996 • M.P.E. Parren 68 (KRIBI, WAG), About 7 km NE of Ebom. Plot 4, subplot 53, tree 9, coordinates: X = 7.5, Y = 8.5 m; 3°07'N, 10°45'E; alt. 500 m; Aug. 1996 • R.G. Letouzey 15272 (P, YA), Piste Meyo Ntem-Evouzok, 75 Km W Ambam, entre 1er et 3e bras du Ntem; 2°16'48"N, 10°31'12"E; 28 Nov. 1979 • T.L.P. Couvreur 1157 (K, MPU, P, WAG, YA), on road Lolodorf-Bipindi, about half way, near Mbiguiligui village (Mbikiliki); 3°09'41.51"N, 10°31'52.18"E; alt. 250 m; 26 Feb. 2018 • T.L.P. Couvreur 486 (YA), 15 km east from Lélé village; 2°16'17.2"N, 13°17'30.35"E; alt. 578 m; 09 Sep. 2013 – South-West Region • A. Binuyo FHI35564 (FHI, K, WAG), Kumba Distr. Along the path from Pete to Bopo at the right handside of the road in Southern Bakundu Forest Reserve; 4°31'N, 9°22'E; 23 Feb. 1956 • A.H. Gentry 62407 (MO), Banyong, Batanga area, between Awong and Banyu, ca 15 km W of Manyemen. TRANSECT 2; 5°00'N, 9°10'E; alt. 420 m; 02 May. 1988 • B. Sonké 5610 (MO); Meme, Nguti, Sanctuaire Mbanyang Mbo; 5°21'24"N, 9°32'57"E; alt. 239 m; 14 Apr. 2011 • C.F.A. Onochie FHI30860 (FHI, K), S.Bakundu, S. Bakundu F.R., between Bombe Rest House and Mbalange; 4°27'N, 9°28'E; 19 Mar. 1953 • D.W. Thomas 3322 (K, MO, P, YA), North-eastern corner of Korup National Park; near Baro village; 5°16'N, 9°11'E; alt. 200 m; 24 Mar. 1984 • D.W. Thomas 4549 (MO, YA), Takamanda Forest Reserve; 6°14'N, 9°19'E; alt. 170 m; 21 Mar. 1985 • D.W. Thomas 5965 (K, MO, WAG, YA), 30 km W of Kumba on Mbonge Road; 4°31'N, 9°22'E; alt. 50 m; 26 Mar. 1986 • D.W. Thomas 6090 (P, YA); Meme, from the vicinity of Lake Barombi, Kumba; 4°39'N, 9°24'E; alt. 300 m; Apr. 1986 • D.W. Thomas 7499 (MO, P, WAG, YA); Ndian, Between Baro and Ikenge villages, along foot path, in the Korup National Park; 5°15'N, 9°00'E; alt. 250 m; 01 Apr. 1988 • G.K. Gottsberger 210307/12 (ULM, WAG), c. 20 m from Banyang Mbo Research Station; 5°08'N, 9°30'E; 21 Mar. 2007 • J. Dundas FHI13898 (K), Southern Bakundu Forest Reserve; 4°30'N, 9°30'E; 19 Feb. 1946 • J. Nemba 64 (K, MO, P, WAG, YA, YA), Bolo Forest, 5 kms W of Kumba – Mamfe road near Konye; 4°55'N, 9°36'E; alt. 300 m; 25 Mar. 1986 • J. Olorunfemi FHI30561 (FHI, K), Mungo River F.R., Kumba Distr.: Mumbo – Southern Bakossi; 4°50'N, 9°20'E; 09 May. 1951 • M.R. Cheek 9337 (K, YA), Mungo River F.R., Mungo river forest reserve. c. 1 Km East of bridge, Chained road to S; 4°44'N, 9°33'E; alt. 200 m; 24 Oct. 1998 • R.G. Letouzey 13673 (P, YA), entre Babong et Okurikang, 35 km WSW. Mamfe (feuille IGN 1/200 000 Mamfe); 5°37'02.16"N, 9°02'22.7"E; 29 May. 1994 • T.L.P. Couvreur 1013 (MPU, WAG, YA), Bayang Mbo Wildlife Sanctuary, after Mbu river; 5°21'19.19"N, 9°30'01.69"E; alt. 242 m; 26 Mar. 2016 • T.L.P. Couvreur 980 (WAG, YA), on top of hill, near Small Ekombe village, 3 km after Kumba on road to Ekondo Titi town; 4°37'22.48"N, 9°22'37.12"E; alt. 615 m; 13 Jan. 2016 • T.L.P. Couvreur 999 (MPU, WAG, YA), Bayang Mbo Wildlife Sanctuary, after Mbu river; 5°21'04.06"N, 9°30'01.78"E; alt. 251 m; 25 Mar. 2016. Gabon – Ogooué-Ivindo • J. Florence 1005 (P), Station d'Ipassa, 10 km S de Makokou; 0°30'N, 12°45'E; alt. 500 m; 17 Apr. 1978 • N. Hallé 548 (P), Bélinga, mines de fer; 1°08'N, 13°12'E; alt. 700 m; 16 Aug. 1966. Ghana – Ashanti Region • C. Vigne 1611 (K, P), Kwahu Prasu; 6°37'10.2"N, 0°54'27.72"E; Feb. 1929 • H. Abbin GC43343 (K), Tano Ofin Forest Reserve, Tano Ofni F.R; 6°40'N, 2°10'W; 16 Aug. 1972 – Eastern Region • F.R. Irvine 3016 (K), Bunsu; 6°16'21.42"N, 0°27'52.93"E; May. 1938 – Western Region • A.A. Enti FE-2169 (B), Neung Forest Reserve, Takoradi-Tarkwa Road; 5°11'07.08"N, 1°58'23.52"W; May. 1982 • A.A. Enti FH6705 (K, P, WAG), Enchi Distr., Enchi – Nyankamam; 5°49'N, 2°49'W; May. 1957 • M.C. Merello 1385 (MO), Bia National Park, Bia National Forest and Production Reserve. Secondary logging roads west from MIM Timber Company Camp; 6°24'15"N, 3°02'30"W; alt. 140 m; 04 Mar. 1996. Ivory Coast – Adzopé • L. Aké Assi 11501 (G), Forêt d'Abongoua; 6°15'N, 3°32'W; 19 Mar. 1971 – Divo • C. Chatelain 679 (G), Forêt de l'IRCC de Divo; 5°47'N, 5°17'W; 19 Dec. 1990 • L. Aké Assi 8469 (G), Forêt de Mopri; 5°46'10.5"N, 4°56'34.87"W; 22 Jan. 1966 – Tiassalé • L. Aké Assi 4290 (G, P), Forêt d'Amitioro; 5°52'50.13"N, 4°52'46.45"W; 07 May. 1957. Nigeria – Cross River State • B.O. Daramola 641 (MO), Ikom Forest Reserve; 5°58'N, 8°42'E; 06 May. 1995 • J.O. Ariwaodo 447 (FHI, WAG); Ikom District, Efraya; 5°53'N, 8°42'E; 25 Mar. 1977 • M.G. Latilo FHI43924 (K); Ikom District, Cross River North Forest Res., between miles 156 & 157 on Ikom-Namfe road; 5°53'08.13"N, 8°47'51.41"E; 10 May. 1961 • P.A. Talbot 123 (K), Oban; 5°13'23.28"N, 8°33'06.9"E; 1911 • P.A. Talbot s.n (BM); Calabar, Oban; 5°13'23.28"N, 8°33'06.9"E; 1912 • P.P.C. van Meer 1490 (WAG), Ekinta River Forest Reserve, Northern part. Near and between pillar 18 and 19; 4°58'N, 8°35'E; 26 Apr. 1971 • P.P.C. van Meer 1664 (WAG); Ikom District, Cross River North Forest Reserve. Compt 1 and 2. 15 km SE of Ikom; 5°52'N, 8°46'E; 18 May. 1971 • P.P.C. van Meer 1736 (WAG); Ikom District, Cross River North Forest Reserve. Compt 1 and 2. 15 km SE of Ikom; 5°52'N, 8°46'E; 20 May. 1971 • R.W.J. Keay FHI28683 (K, P); Ikom District, Cross River North Forest Res.; 5°50'N, 8°50'E; 19 Jan. 1951 – Nassarawa State • A.P.D. Jones FHI16965 (P), Khaya HF nr. Onda enclave; 8°30'N, 7°48'E; 18 Feb. 1946 – Oyo State • C.F.A. Onochie FHI31539 (K); Ibadan District, c. 14 miles south of Ibadan on the Ijebu-Ode road; 7°11'00.66"N, 3°58'13.92"E; 19 Mar. 1958 • D.P. Stanfield FHI44949 (K, MO); Ibadan District, Gambari Forest Reserve; 7°07'59.88"N, 3°49'59.88"E; alt. 80 m; 11 Mar. 1964 • R.W.J. Keay FHI22812 (K); Ibadan District, Ibadan Forest Reserve, c. 22 miles south of Ibadan. School Enumeration area 1948; 7°03'02.43"N, 3°54'20.56"E; 22 Apr. 1948.
Uvariodendron citriodorum , stat. nov.
≡ Uvariodendron molundense var. citrata Le Thomas, Fl. Gabon No. 16, 283 (1969). Type. Gabon – Ogooué-Ivindo • N. Hallé 525 (holotype: P! (P00363400), sheet here designated, isotypes: K! (K000198798, K000198799), MO! (MO216993, MO216994), P! (P00363398), WAG! (WAG0065739)), Bélinga, mines de fer, sommet de Belvédère; 1°08'N, 13°12'E; alt. 800 m; 15 Aug. 1966.
Description
Tree to shrub 4–5 m tall, D.B.H. unknown; young branches glabrous, old branches glabrous. Leaves with strong lemon smell when crushed. Petiole 6–10 mm long, 1.5–3.5 mm wide, sparsely pubescent to glabrous. Leaf lamina (183) 250–337 mm long, 72–97 mm wide, length:width ratio 2.3–4.2, elliptic to oblong, coriaceous, base acute to rounded (sometimes minutely decurrent at the very base, and decurrent on young leaves), apex attenuate, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 12–18 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on branches or axillary, composed of 1 sessile flower. Flowers bisexual, buds globose, sessile, 5–6 mm high, 5–7 mm in diameter, velutinous. Mature flower unknown, measures taken from flower buds or fruits. Bracts 1 to 4, upper bract ca. 5 mm long, ca. 7 mm wide, broadly ovate, semi clasping the pedicel, velutinous outside, glabrous inside. Sepals 3, ca. 7 mm long, ca. 6 mm wide (measures taken from bud), imbricate, velutinous outside, glabrous inside, color unknown. Outer petals 3, length, shape, indumentum and color unknown. Inner petals 3, length, shape, indumentum and color unknown. Stamens number unknown, ca. 1 mm long, ca. 0.2 mm wide, anthers linear, connective prolongation truncate. Carpels 10 to 20, ca. 2 mm long, ca. 0.4 mm wide, sparsely pubescent, free; stigma 0.5 mm long, 0.5 mm wide, pubescent. Fruiting pedicel 10–12 mm long, 2–5 mm in diameter, pubescent to glabrous. Monocarps 5 to 20, 25–40 mm long, 9–20 mm wide, length:width ratio 1.9–2.9, cylindrical, oblong, straight, truncate or rounded at apex, pubescent to glabrate, pale greyish green to red; sessile to shortly stipitate, stipe 0–5 mm long, 3–5 mm wide, pubescent to glabrous. Seeds 9–14 per monocarp, biseriate, 13 to 15 mm long, ca. 11 mm wide, semicircular.

Uvariodendron citriodorum (Le Thomas) Dagallier & Couvreur A entire specimen sheet with branch, leaves and flower buds B flower bud showing bracts, semi-side view C fruit with monocarps, some monocarps transversally cut, top view D fruit with monocarps, some monocarps transversally cut, side view. A, B Hallé 525 (type) C, D Sosef 2219. Photos Léo-Paul Dagallier.
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region. Known from one area in Gabon: Bélinga, mines de fer, and one locality in Republic of the Congo: Les Saras.
Habitat and ecology
Mature rain forests on rocky soils. Altitude: 800–950 m a.s.l.
Phenology
Fruits collected in July and November.
Vernacular names
Gabon: ‘Bombamba’ in Bakota (Hallé 2896).
Etymology
The specific epithet refers to the strong lemon smell of the plant. The epithet ‘citrata’ would prevail as it is the one given by Le Thomas for the variety. However, this specific epithet is already taken by Uvariopsis citrata Couvreur & Niangadouma, from the genus Uvariopsis, which is closely related to Uvariodendron. Following the recommendation 23.A.3.h from

Notes
This species differs from the other species, and particularly from Ud. molundense, in having leaves emitting a strong lemon scent when fresh material is crushed. Apart from this character, this species is morphologically very similar to Ud. molundense. However, as Ud. citriodorum and Ud. molundense are not monophyletic (Fig.
Preliminary conservation status
This species is only known from one locality in Gabon and one locality in the Republic of the Congo. The EOO of the species is estimated at 2,256 km2 and its AOO at 20 km2. The unique occurrence in the Republic of the Congo dates back more than 30 years and the occurrences in Gabon are situated in Bélinga. Although the project of exploitation of the Bélinga iron ore deposits has been canceled, this locality is still threatened by a possible future exploitation of the ore. Following IUCN criterion B, it is thus assigned a preliminary conservation status of Endangered EN B1a(i,ii,iii,iv)+2a(i,ii,iii,iv).
Additional specimens examined
Gabon – Ogooué-Ivindo • M.S.M. Sosef 2219 (BR, K, LBV, MO, WAG), Bélinga, Mines de Fer, 4 km on the road to Mvadi; 1°05'N, 13°12'E; alt. 900 m; 05 Nov. 2005 • N. Hallé 2896 (P), Bélinga; 1°05'N, 13°08'E; alt. 900 m; 29 Oct. 1964 • N. Hallé 3082 (P), Bélinga; 1°05'N, 13°08'E; alt. 950 m; 07 Nov. 1964 • N. Hallé 13 (P), Bélinga, mines de fer, route du Belvédère; 1°05'N, 13°12'E; alt. 950 m; 08 Jul. 1966 • N. Hallé (P), Bélinga, mines de fer, route du pt. B3; 1°06'N, 13°12'E; 25 Jul. 1966 • N. Hallé (K, MO, P), Bélinga, mines de fer; 1°08'N, 13°12'E; alt. 950 m; 16 Jul. 1966. Republic of the Congo – Kouilou • H. de Foresta 1759 (P), Les Saras, piste Cofibois, environs de la plantation Coobama de 1987; 4°22'S, 12°22'E; 25 Nov. 1988.
Uvariodendron connivens
≡ Uvaria connivens Benth., Trans. Linn. Soc. London 23(3): 465 (1862); Uva connivens Kuntze, Revis. Gen. Pl. 1: 8. (1891). Type. Equatorial Guinea – Bioko Sur • G. Mann 1159 (lectotype: K! (K000198803); isolectotypes: P! (P00362655), K! (K000198804, K000198805)), Fernando Po; 3°30'N, 8°40'E; 1861.
= Uvaria winkleri Diels, Bot. Jahrb. Syst. 38(3): 240 (1907). Type. Cameroon – South-West Region • H. Winkler 1466 (holotype: B (not found, destroyed?)); Moliwe.
= Uvaria megalantha
Diels, Bot. Jahrb. Syst. 39(3–4): 472 (1907). Type. Cameroon – South Region • G.A. Zenker 3204 (holotype: B (not found, destroyed?); lectotype: WAG! (WAG0057972), designated by
Description
Tree 3–20 m tall, D.B.H. 2–25 cm; young branches glabrous, old branches glabrous. Petiole 5–21 mm long, 2–6 mm wide, glabrous. Leaf lamina 254–636 mm long, 66–177 mm wide, length:width ratio 2.3–6.2, elliptic to oblong to obovate, coriaceous, base acute to rounded, sometimes truncate or subcordate, apex acute to acuminate, acumen 5–24 mm long; surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 16–28 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and branches or axillary, composed of 1 flower. Flower pedicel (5) 10–40 mm long, 1.4–4 mm, increasing toward the apex up to 10 mm in diameter, pubescent to glabrous. Flowers bisexual, buds ovoid to globose, pedicellate, 9–22 mm high, 8–30 mm in diameter, pubescent. Bracts 1 to 6, upper bract 4–13 mm long, 6–17 mm wide, ovate, appressed, enclosing the bud, pubescent outside, glabrous inside. Sepals 3, 5–14 mm long, 7–17 mm wide, imbricate to fused at base, puberulent outside, glabrous inside, dull green. Outer petals 3, 14–32 mm long, 12–29 mm wide, length:width ratio 0.95–1.5, broadly ovate to ovate, pubescent to puberulent outside, puberulent to glabrous inside, wine red (cream in immature flowers) outside, wine red (cream in immature flowers) inside. Inner petals 3, 9.5–30 mm long, 8–23 mm wide, length:width ratio 0.9–1.9, broadly ovate to ovate, pubescent to puberulent outside, puberulent to glabrous inside, wine red (cream in immature flowers) outside, wine red (cream in immature flowers) inside. Stamens 1900 to 2500, 2–3.8 mm long, 0.3–0.5 mm wide, anthers linear, connective prolongation truncate. Carpels 5 to 33, 2–6.5 mm long, 0.8–1.5 mm wide, pubescent, free; stigma 0–2 mm long, 1–1.8 mm wide, coiled, pubescent, covered with an exudate at anthesis. Fruiting pedicel 13–31 mm long, 3–8 mm in diameter, glabrous. Monocarps 1 to 10, 22–55 mm long, 17–32 mm wide, length:width ratio 1.3–2, cylindrical to ovoid, longitudinally ridged, sparsely pubescent to glabrous, red to orange when ripe, green when immature; stipe 0–8 mm long, 3–10 mm wide, slightly pubescent to glabrous. Seeds 8–23 per monocarp, biseriate, 13 to 29 mm long, 3–14 mm wide, semicircular.

Uvariodendron connivens (Benth.) R.E.Fr. A young branch with leaves, upper side B detail of petiole and base of leaf, upper side C fruiting material with unripe monocarp, side view D flower, top view E flower, side view F flower bud, top view G flower bud, bottom view H flower, one sepal removed, semi-bottom view. A, B Couvreur 383 C, F, G Couvreur 620 D, E Couvreur 1016 H Couvreur 1051. Photos Thomas Couvreur.

Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Cameroon, Equatorial Guinea (Bioko Island) and Nigeria.
Habitat and ecology
Lowland and premontane mature and old secondary rain forests, sometimes in swamp forests. Altitude: 50–1000 m a.sl.
Phenology
Flowers and fruits collected all year.
Vernacular names
Cameroon: ‘Ikeinju’ in Bakweri (Mbani 14).
Uses
The young leaves are eaten (Cheek 5180) and the fruits are used for cough and dye (Mbani 14).
Notes
This species is easily differentiated from the other species by its petals being wine red on both sides in mature flowers. Apart from this character, Ud. connivens resemble Ud. calophyllum and Ud. fuscum in having great leaves (more than 25 cm long). It can be distinguished from these species by being glabrous when young and old, and by having a distinct flower pedicel 10–40 mm long, whereas Ud. calophyllum and Ud. fuscum flowers are subsessile (flower pedicel 10–15 mm maximum in Ud. fuscum var. magnificum).
Preliminary conservation status
This species has been assessed previously as Near Threatened NT but the assessment needs to be updated (
Additional specimens examined
Cameroon – Littoral • T.L.P. Couvreur 620 (MPU, YA), Ebo Wildlife Reserve, Djuma permanent camp. On Djuma-Djuma trail; 4°20'23.59"N, 10°14'41.58"E; alt. 335 m; 14 Feb. 2014 – South Region • G.A. Zenker 2624 (B, BM, G, K, L, M, MA, P, P, WAG), Bipindi; 3°05'N, 10°25'E; 1903 • G.A. Zenker 3401 (BM, G, K), Bipinde; 3°05'N, 10°25'E; 1907 • G.A. Zenker 3487 (BM, G, K), Bipinde; 3°05'N, 10°25'E; 1908 • G.A. Zenker 358 (B, G, M, P, P, U, WAG), Mimfia; 3°04'N, 10°23'E; Sep. 1913 • G.A. Zenker 3845 (BM, K), Bipindi; 3°05'N, 10°25'E; 1909 • J.J. Bos 5412 (P, WAG), 6 km S. of Kribi, 2–4 km E. of Grand Batanga road; 2°53'N, 9°55'E; 26 Sep. 1969 • T.L.P. Couvreur 383 (MPU, WAG, YA); Océan, Campo Ma an National Park, 5 km after main entrance; 2°21'19.48"N, 10°15'33.59"E; alt. 300 m; 15 Feb. 2012 • T.L.P. Couvreur 484 (MPU, YA), 15 km east from Lélé village; 2°16'39.39"N, 13°17'37.19"E; alt. 549 m; 09 Sep. 2013 • T.L.P. Couvreur 706 (MPU, WAG, YA), Campo Ma'an National Park, 11 km on trail from Ebinanemeyong village, on road, 7 km from Nyabessan to Campo town; 2°28'25.42"N, 10°20'39.1"E; 14 Feb. 2015 – South-West Region • B.-A. Nkongmeneck 959 (YA); Fako, Idenau, Mt Cameroun, versabt de Idenao. Feuille IGN 1/200 000 Buea/Douala; 4°12'N, 9°05'E; alt. 300 m; 23 Feb. 1985 • C.F. Tekwe 87 (K, SCA, YA), Isobi, above Isobe; 4°10'N, 9°00'E; alt. 40 m; 10 Jun. 1992 • D. Kenfack 1507 (MO), Mokoko; 4°27'N, 9°04'00.12"E; 24 Apr. 2001 • D. Kenfack 879 (MO); Ndian, Korup National Park, 11 km from Mundemba, along Fabe road; 5°05'03.43"N, 9°32'20.63"E; alt. 9 m; 11 Jul. 1997 • D.W. Thomas 2264 (K, MO, P, WAG, YA); Ndian, Map # NB 32 IV Buea-Douala. South Korup Reserve; 4°55'N, 8°50'E; alt. 50 m; Jul. 1983 • D.W. Thomas 4447 (K, MO, P, YA), forest and meadows on the gently sloping side of Mt Cameroun above small Koto village; 4°18'N, 9°06'E; alt. 550 m; 06 Mar. 1985 • D.W. Thomas 5533 (P, YA), forest between Kindonge and small Ekombe, Southern Bakundu Forest Reserve; 4°35'N, 9°23'E; alt. 200 m; 10 Feb. 1986 • D.W. Thomas 5537 (P, YA), at Bonenza, 2 km N of Limbe-Idenao road; 4°03'N, 9°05'E; alt. 300 m; 10 Feb. 1986 • D.W. Thomas 6928 (P, YA), footpath toward Matene from Mbilishe; 6°14'N, 9°26'E; alt. 400 m; 1987 • D.W. Thomas 7961 (MO, P, YA), Steep hillside south of Esukutang village; 5°23'N, 9°00'E; alt. 300 m; 25 May. 1988 • D.W. Thomas 9875 (K, SCA, YA), West of the Onge River and ridges on ‘Thump Mount'; 4°20'N, 8°57'E; alt. 200 m; 09 Nov. 1993 • F. Nguembock 76 (K); Fako, Mabeta-Moliwe Reserve; 3°58'N, 9°14'E; 10 Mar. 1992 • G. Mann 763 (BM, K), Ambas Bay; 4°01'N, 9°12'E; Feb. 1861 • G.K. Gottsberger 130307/21 (ULM, WAG), c. 300 m from Banyang Mbo Research Station; 5°08'N, 9°30'E; 13 Mar. 2007 • G.P. Tchouto Mbatchou 685 (K, SCA, YA), Bomana secondary forest. Transect OA, Plot OA0Z; 4°15'N, 9°01'E; alt. 200 m; 05 Oct. 1993 • I. von Rege 87 (K, SCA, YA); Fako, Mabeta, 6 km SE Limbe SBL; 3°59'N, 9°17'E; alt. 60 m; 11 Aug. 1993 • J. Bongyu 73 (K); Fako, Mabeta-Moliwe Reserve; 3°58'N, 9°14'E; 06 Apr. 1992 • J. Nemba 550 (MO, P, YA), Secondary growth and old growth forest along Kumba-Mamfe road at mile 14 Lkiliwindi; 4°44'N, 9°29'E; alt. 200 m; 15 Jun. 1987 • J. Nemba 56 (L, P, U, YA), 5 kms west of Kumba-Mamfe road near Konye; 4°55'N, 9°36'E; alt. 300 m; 25 Mar. 1986 • J.-P. Ghogue 1551 (YA), Bimbia Bonadikombo (former Mabeta Moliwe), 18 km SE Limbe; 3°59'36"N, 9°15'43"E; alt. 50 m; 10 Apr. 2003 • J.F. Villiers 2483 (P, YA), 4 km E Bomana, 34 NW limbé; 4°12'55.22"N, 9°06'11.21"E; 14 Dec. 1984 • J.I. Wheatley 194 (K, SCA, YA); Fako, Mabeta-Moliwe TC 10; 4°01'N, 9°16'E; alt. 50 m; 20 Apr. 1992 • J.I. Wheatley 326 (K, SCA, YA); Fako, Mabeta-Moliwe TD 5835 m; 4°01'N, 9°16'E; alt. 100 m; 24 Jun. 1992 • J.J. Wieringa 45 (WAG); Fako, Limbe, Bakingini, forest above ‘mile 11 village'; 4°04'10"N, 9° 03'50"E; alt. 160 m; 20 Jan. 1994 • J.J. Wieringa 5839 (BR, FHO, MO, WAG), Bakingini, at edge of plantation area above Mile 11; 4°04'10.8"N, 9°03'27.6"E; alt. 150 m; 06 Mar. 2007 • J.M. Dalziel 8243 (K), Buea1, Buea to Mayuko; 4°09'N, 9°14'E; 13 Feb. 1927 • J.M. Mbani 14 (K, SCA, YA), Njonji; 4°08'11.04"N, 8°59'22.56"E; alt. 100 m; 13 Feb. 1992 • J.M. Mbani 382 (K, SCA, YA), Ekumbe Mofako, Plot M19; 4°28'N, 9°04'E; 21 May. 1994 • J.P. Watts 245 (K, YA), Moliwe, Makota River watershed.TD 5832 m; 4°00'N, 9°15'E; alt. 100 m; 29 Apr. 1992 • J.P. Watts 394 (K, SCA, YA), c.5 km South East of Moliwe, TF +4000 m; 4°02'N, 9°17'E; alt. 100 m; 09 Jun. 1992 • J.P. Watts 782 (K, SCA, YA), forest to the West of Onge River, about 4 km West of Liwenyi village (c 14 km North of Idenau); 4°23'N, 8°59'E; alt. 260 m; 28 Oct. 1993 • M. Etuge 156 (MA, MO, P, WAG, YA), Bakolle Bakossi, on Kumba – Mamfe road; 5°01'N, 9°40'E; alt. 350 m; 24 May. 1986 • M. Etuge 2390 (K, YA), Max's trail, Nyasoso; 4°49'39.72"N, 9°40'51.96"E; alt. 1100 m; 24 Jun. 1996 • M. Etuge 2396 (K, YA), Max's trail, Nyasoso; 4°49'39.72"N, 9°40'51.96"E; alt. 1100 m; 24 Jun. 1996 • M. Etuge 6506 (YA), Mungo River F.R., Mungo F.R; 4°44'17"N, 9°33'38"E; 22 Feb. 2006 • M.R. Cheek 5180 (K, P, SCA, YA), Liwenyi, Low altitude forest on the West bank of the Onge river above the first set of rapids, that is about 1–2 hours walk inland from Enyenge. Grid ref. and alt. approx. Local names and uses from Clement Offu (Enyenge); 4°17'N, 8°58'E; alt. 50 m; 28 Oct. 1993 • M.R. Cheek 5462 (K, SCA, YA), Low altitude forest above oil palm plantation. Reached after c. 40 minutes walk N then E from Njonji. Hunters path to ‘Lake Njonji'. Little farming, but many gaps and fallen trees at low altitude, c. 150–300 m, crossing and running alongside substantial seasonal stream. At c. 400 m, Hypselodelphyus-Aframomum thicket dominates; 4°08'N, 9°01'E; alt. 300 m; 18 Nov. 1993 • M.R. Cheek 8164 (K, YA); Ndian, Korup National Park, Ekundu Kundu, Transect 10, c. 1100 m; 5°08'N, 8°55'E; alt. 170 m; 25 Apr. 1996 • N. Ndam 1076 (K, SCA), Bonjare, Plot 09; 4°26'N, 9°01'E; alt. 220 m; 30 Apr. 1994 • N. Ndam 1118 (K, SCA), Bonjare, Plot 10; 4°26'N, 9°01'E; alt. 270 m; 01 May. 1994 • N. Ndam 708 (K, YA), Bomana-Koto Rd c 500 m Bearing 305deg towards Onge river 3 hr walk from the rd; 4°13'N, 9°04'E; alt. 400 m; 18 Oct. 1993 • P. Nkeng 37 (K, SCA, YA), Etome; 4°03'N, 9°07'E; alt. 300 m; 02 Mar. 1992 • R.G. Letouzey 15175 (P, YA); Ndian, Rivières Mosongosele et de Ndian depuis Mosongosele jusqu'à l'entrée amont de la mangroce, env. 20 km au SW de Mundemba (feuille IGN 1/200 000 Buea-Douala); 4°49'27.3"N, 8°45'15.72"E; 13 Jun. 1976 – R.G. Letouzey 15177 (MO, P, P, WAG, YA), Au SW de Mosongosele, 20 km WSW Mendumba; 4°54'N, 8°45'E; 14 Jun. 1976 • S. Cable 2329 (K, YA); Ndian, Korup National Park, Ekundu Kundu, path to Esoki about 4 km from Ekundu-Kundu (as on 27th); 5°03'16.8"N, 8°56'58.5"E; alt. 350 m; 29 Apr. 1996 • S. Cable 611 (K, SCA, YA), Dikulu, Coastal lowland rain forest along Mangrove stream; 3°59'N, 9°14'E; alt. 50 m; 17 Dec. 1993 • S.N. Ekema 1078 (K, YA), Mokoko Forest Reserve, Boa/Likinge; 4°24'N, 9°00'E; alt. 150 m; 31 May. 1994 • T.C.H. Sunderland 1264 (K, SCA, YA), Mabeta-Moliwe: TB 6000 m; 4°03'N, 9°16'E; alt. 40 m; 22 Apr. 1992 • T.C.H. Sunderland 1536 (K, K, SCA, YA), Nyasoso, Mt Kupe: Max's Trail; 4°48'05.04"N, 9°42'29.16"E; alt. 1600 m; 09 Jul. 1992 • T.D. Maitland 537 (K, P), Buea area, at Balifamba; 4°10'N, 9°18'E; alt. 731 m; 1929 • T.L.P. Couvreur 1016 (MPU, WAG, YA), Bayang Mbo Wildlife Sanctuary, after Mbu river; 5°21'26.28"N, 9°30'05.88"E; alt. 253 m; 26 Mar. 2016 • T.L.P. Couvreur 1051 (MPU, WAG, YA), Mt Cameroon National Park, on the Bomona trail, behind Bomona village, 10 km NW from Idenau; 4°17'48.27"N, 9°04'43.77"E; alt. 690 m; 03 Apr. 2016 • W.G. Gosline 235 (K, YA), Kupe Village, trail to Kupe rock saddle; 4°47'10"N, 9°41'30"E; alt. 950 m; 28 Nov. 1999 • W.J. Baker 294 (K, SCA, YA); Fako, Mabeta, 6 km SE Limbe SBL; 3°59'N, 9°17'E; 10 Aug. 1993 – Unknown major area • W.G. Gosline 209 (K, WAG, YA), Meme Division, Mahole-Bintulu road; 4°47'30"N, 9°36'12"E; alt. 300 m; 24 Nov. 1999. Equatorial Guinea – Bioko Norte • M.G. Carvalho 4220 (K, MA), BIOCO: Malabo – Cupapa, km 22–23, margenes del rio Ejoa, 32NMK9005, 200 m; 3°43'12"N, 8°51'36"E; 06 Jul. 1989 – Bioko Sur • W.R.Q. Luke 11955 (EA, K, MA), Moaba – Moka trail, Biadyi River Camp Pt 140; 3°17'06.4"N, 8°38'17.81"E; alt. 650 m; 16 Mar. 2007 • W.R.Q. Luke 13184 (K, MA), Moraka pt 340 to 342; 3°16'13.8"N, 8°28'29.64"E; alt. 3 m; 28 Jan. 2009. Nigeria – Cross River State • C.F.A. Onochie FHI36080X (K); Calabar, Oban Group Forest Reserve, between Akor and Orem; 5°36'N, 8°35'E; 21 Jan. 1957 • E.U. Ujor FHI31636 (K); Calabar, Ikot Ewa; 4°57'24.84"N, 8°38'44.88"E; 05 Jul. 1952 • H.D. Onyeachusim FHI54055 (K); Calabar, between miles 57–58 Osomba village on Calabar-Mamfe road; 5°27'23.32"N, 8°39'55.34"E; 21 Feb. 1964 • M.C. Ejiofor FHI21898 (K); Calabar, Oban Group Forest Reserve; 5°36'N, 8°35'E; 07 May. 1952 • M.G. Latilo 23 (K); Calabar, Akamkpa Rubber Estate. Calabar River Division; 5°18'N, 8°20'E; 21 Mar. 1959 • M.G. Latilo 32 (K, MO); Calabar, Akamkpa Rubber Estate. Dukwe felling area. Calabar River Division; 5°18'N, 8°20'E; 23 Mar. 1959 • P.A. Talbot 1254 (BM, K), Oban; 5°13'23.28"N, 8°33'06.9"E; 1911 • P.A. Talbot 158 (BM); Calabar, Oban; 5°19'N, 8°34'E; 1911 • P.A. Talbot 402 (BM), Oban; 5°13'23.28"N, 8°33'06.9"E; 1911 • P.A. Talbot 404 (BM, K), Oban; 5°13'23.28"N, 8°33'06.9"E; 1911 • P.A. Talbot 433 (BM), Oban; 5°13'23.28"N, 8°33'06.9"E; 1911 • P.A. Talbot 434 (BM), Oban; 5°13'23.28"N, 8°33'06.9"E; 1911 • P.A. Talbot 82 (BM, K), Oban; 5°13'23.28"N, 8°33'06.9"E; 1911 • P.A. Talbot s.n (BM); Calabar, Oban; 5°13'23.28"N, 8°33'06.9"E; 1912 • P.P.C. van Meer 1412 (WAG); Calabar, Oban Group Forest Reserve, East Block; 5°31'N, 8°41'E; alt. 200 m; 21 Apr. 1971 • P.P.C. van Meer 1430 (U); Calabar, Oban Group Forest Reserve, East Block; 5°31'N, 8°41'E; alt. 200 m; 22 Apr. 1971 • P.P.C. van Meer 1430 (WAG); Calabar, Oban Group Forest Reserve, East Block; 5°31'N, 8°41'E; alt. 200 m; 22 Apr. 1971 • R.W.J. Keay FHI28191 (K); Ikom District, Afi River Forest Reserve, near Aboabam, forest by river Nkem; 6°11'52.16"N, 8°58'40.48"E; 09 Dec. 1950.
Uvariodendron dzomboense
Type
Kenya – Coast • S.A. Robertson MDE207 (holotype: K! (no barcode); isotypes: EA!, MO!, WAG! (WAG0065798)), Kwale District – Dzombo Hill; 4°26'S, 39°13'E; alt. 300 m; 07 Feb. 1989.
Description
Tree 4–7 m tall, D.B.H. unknown; young branches sparsely pubescent to glabrous, old branches glabrous; leaf bud ‘eragrostiform’, composed of 5, distichous, longitudinally folded, velutinous scales. Leaves with margin slightly revolute. Petiole 3–4 mm long, 1–1.5 mm wide, sparsely pubescent to glabrous. Leaf lamina 65–132 mm long, 20–45 mm wide, length:width ratio 2.9–3.6, elliptic to narrowly elliptic, coriaceous, base acute to slightly decurrent, apex attenuate, surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, slightly pubescent to glabrous below; secondary veins 12–13 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and branches, composed of 1 flower. Flower pedicel 8–30 mm long, 2–2.5 mm in diameter, densely pubescent. Flowers bisexual, buds globose, sessile, ca. 4 mm high, ca. 4.5 mm in diameter, sparsely pubescent. Bracts 6 at base of the pedicel in flower bud, generally the uppermost remaining towards the lower half of the pedicel on mature flower, 5–6 mm long, 5–8 mm wide, ovate, pubescent to shortly pubescent outside, glabrous inside. Sepals 3, 5–7 mm long, 4.5–7 mm wide, fused at base, pubescent to shortly pubescent outside, glabrous inside, color unknown. Outer petals 3, ca. 16 mm long, ca. 9 mm wide, length:width ratio ca. 1.8, ovate, shortly velutinous outside, glabrous inside, color unknown. Inner petals 3, ca. 18 mm long, ca. 8 mm wide, length:width ratio ca. 2.3, obovate, shortly velutinous outside, glabrous inside, color unknown. Stamens 700 to 1000, 2 mm long, 0.5 mm wide, anthers linear, connective prolongation truncate. Carpels 50 to 75, ca. 2 mm long, 1–1.5 mm wide, densely pubescent, free; stigma unknown. Fruiting pedicel ca. 14 mm long, ca. 4 mm in diameter, pubescent. Monocarps (unripe?) ca. 35, ca. 15 mm long, ca. 10 mm wide, length:width ratio ca 1.5, ovoid, densely pubescent, golden brown, sessile. Seeds (unripe?) ca. 5 per monocarp, uniseriate, ca. 4.5 mm long, ca. 10 mm wide.

Uvariodendron dzomboense Dagallier, Q.Luke & Couvreur A leaf, upper side B petiole and base of leaf, upper side C flower bud showing bracts, side view D flower, two outer petals and three outer petals removed E fruit with young monocarps. A–C Luke 7443 D, E Robertson MDE 207. Photos Léo-Paul Dagallier.
Distribution
Endemic to Somalia-Masai Region. Only known from one locality in Kenya: Dzombo Hill.
Habitat and ecology
Moist semi-deciduous forest. Soil: igneous intrusion. Altitude: 270–300 m a.s.l.
Phenology
Flowers collected in January and June. Fruits collected in February.
Notes
This species differs from the other Uvariodendron species by the combination of small (i.e. less than 150 mm long) narrowly elliptic to elliptic leaves and 50–75 densely pubescent carpels. It differs from Ud. kirkii by its smaller leaves (132 mm maximum versus 210 mm maximum) and higher number of carpels (50–75 versus 7–20).

Preliminary conservation status
Previous work estimated the EOO and AOO of this species to be less than 6 km2, and assigned a preliminary status of Endangered EN B1ab(iii)+2ab(iii) (
Additional specimens examined
Kenya – Coast • W.R.Q. Luke 1654 (EA, K); Kwale District, Dzombo Forest Reserve; 4°25'S, 39°13'E; alt. 270 m; 06 Jan. 1989 • W.R.Q. Luke 2884 (EA, K); Kwale District, Dzombo Forest Reserve; 4°25'S, 39°13'E; alt. 271 m; 04 Oct. 1991 • W.R.Q. Luke 3370 (EA); Kwale District, Dzombo Forest Reserve; 4°25'S, 39°13'E; alt. 272 m; 12 Nov. 1992 • W.R.Q. Luke 7443 (EA); Kwale District, Dzombo; 4°25'S, 39°12'E; alt. 270 m; 28 Jun. 2001.
Uvariodendron fuscum
≡ Uvaria fusca Benth., Trans. Linn. Soc. London 23(3): 466 (1862); Uva fusca Kuntze, Revis. Gen. Pl. 1: 8 (1891). Type. Equatorial Guinea – Bioko Sur – G. Mann 308 (holotype: K! (K000198801); isotype: P! (P00362657)); 3°30'N, 8°40'E; alt. 396 m; 1860.
= Uvariodendron mirabile
R.E.Fr., Acta Horti Berg. 10 : 59 (1930). Type. Cameroon – South-West Region • P.R. Preuss 1378 (lectotype: P! (P00315830), designated by
= Uvaria gigantea Engl.; Uva gigantea Kuntze; Uvariodendron giganteum (Engl.) R.E.Fr.; concerning Uvariodendron fuscum var. giganteum (see details under this variety).
= Uvariodendron magnificum Verdc.; syn. nov. concerning Uvariodendron fuscum var. magnificum (see details under this variety).
Description
Tree 3–15 m tall, D.B.H. 5–35 cm; young branches with long soft hairs producing a whitish appearance quickly falling off to glabrous, old branches glabrous. Petiole 4–35 mm long, 2–8 mm wide, pilose to glabrous. Leaf lamina 160–750 mm long, 43–225 (250) mm wide, length:width ratio (2.1) 2.5–4, narrowly elliptic to elliptic to narrowly obovate, coriaceous, base acute to cuneate to rounded, apex rounded to acuminate, acumen 1–23 mm long; surface above glabrous, surface below pilose to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, pilose to glabrous below; secondary veins 15–33 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and branches, composed of 1–2 (sub)sessile to pedicellate flowers. Flower pedicel 0–15 mm long, 3–6 mm in diameter, velutinous. Flowers bisexual, buds globose, sessile, 6–15 mm high, 6.5–15 mm in diameter, velutinous. Bracts 1 to 6, upper bract 8–35 mm long, 10–50 mm wide, broadly ovate, clasping the pedicel, enclosing the sepals, pubescent outside, glabrous inside. Sepals 3, 11–55 mm long, 13–43 mm wide, ovate, fused at base over 20–50% of their length, velutinous outside, glabrous inside, brown. Outer petals 3, 20–70 mm long, 17–47 mm wide, length:width ratio 1.1–1.7, elliptic to broadly ovate, velutinous outside, glabrous inside, cream to greenish outside, cream with a dark red streak from base up to 75% of the petal length inside. Inner petals 3, 20–53 mm long, 15–38 mm wide, length:width ratio 1.1–2.1, elliptic to broadly obovate to obovate, puberulent outside, glabrous inside, cream outside, dark red to dark purplish red with cream margins inside. Stamens around 2500, 3.5–5 mm long, 0.1–0.5 mm wide, anthers linear, connective prolongation truncate, pale yellow. Carpels 20 to 160, 4–7 mm long, 0.5–2.2 mm wide, velutinous, free; stigma 1–2 mm long, 0.8–1.1 mm wide, coiled, glabrous to velutinous, covered with an exudate at anthesis. Fruiting pedicel 5–18 mm long, 5–8 mm in diameter, pubescent. Monocarps 6 to 80, 20–60 mm long, 11–32 mm wide, length:width ratio 1.7–2.5, cylindrical, curved, acuminate, puberulent, green to brown; sessile to shortly stipitate, stipe 0–4 mm long, 2–5 mm wide, pubescent to glabrate. Seeds 1–16 per monocarp, biseriate, 14 to 23 mm long, 7–11 mm wide, semicircular, orange-brown.

Uvariodendron fuscum var. fuscum A leaf, lower side B apex of young branch with detail of petiole and leaf base, upper side, inset: detail of sparse pubescence on young branch C trunk with flower bud (top) and flower (bottom) D open flower E young fruit with unripe monocarps, one old petal remaining F flower semi-bottom view, showing bracts and sepals G detail of flower after anthesis, note the falling stamens and pollinator insects, inset: detail of Coleoptera full of pollen grains. A, E, G Couvreur 1046 B, C Couvreur 1029 D, F Couvreur 990. Photos Thomas Couvreur.
Distribution
Element of the Lower Guinean Domain and Congolia Domain of the Guineo-Congolian Region and Zambezian Region: Cameroon, Democratic Republic of the Congo, Gabon, Equatorial Guinea (Bioko Island), Nigeria, Uganda.
Habitat and ecology
Lowland and submontane to mountain mature or old secondary rain forests. Altitude: 100–1400 m a.s.l.
Phenology
Flowers collected from January to April. Fruits collected from March to April.
Vernacular names
Cameroon: ‘Limboto’ in Bakweri (van Andel 3761), ‘Obom Ossoé’ in Yaoundé (Biholong 279). Gabon: ‘Inkaca’ in Bakota (Hallé 3156).
Uses
The young leaves are boiled and used for soup (Cheek 5145) and the ripe fruits are eaten (van Andel 3761).
Notes
From now on, the name Ud. fuscum encompasses the synonyms Ud. mirabile, Ud. giganteum and Ud. magnificum. Ud. fuscum consists of a large morphological variation of the leaf and flower size, with var. magnificum having the largest dimensions, followed by var. giganteum and then the type variety being the smallest (Fig.

Uvariodendron fuscum var. giganteum (R.E.Fr.) Dagallier & Couvreur A young branch with leaves, lower side, inset: detail of midrib, note the long soft hairs producing A whitish appearance B young branch, petiole and base of leaf, upper side, note the absence of hairs (fallen off) C flower, top view D flower, side view showing sepals and bracts E young fruit, top view. A Couvreur 1229 B–D Couvreur 1057. Photos Thomas Couvreur.
Preliminary conservation status
This species is widespread, distributed from Nigeria to Uganda. A previous assessment listed it as Near Threatened NT (

Uvariodendron fuscum var. giganteum (R.E.Fr.) Dagallier & Couvreur A leaf B flower bud, side view C flower bud, bracts removed, side view D detail of flower bud, two sepals, one outer and two inner petals removed E outer petal, outer view F inner petal, outer view G stamen H carpels, side view and detail of ovules I longitudinal section of receptacle J floral diagram K fruit, longitudinal sections of monocarps. A–K from Hallé 3156 (as Ud. giganteum). Drawings by Hélène Lamourdedieu, modified from

Uvariodendron fuscum var. magnificum (Verdc.) Dagallier & Couvreur A young branch with leaves and apical bud B leaf, upper side C longitudinal section of flower bud D flower, semi-side view E carpel with carpel wall partially removed to show arrangement of ovules, side view F young fruit, side view G monocarp, side view H seed, side view. A–H from Okodi in Hamilton 696 (as Ud. magnificum). Drawings by Mary Griesrson, modified from
Uvariodendron fuscum var. fuscum
Description
Young branches sparsely pubescent to glabrous. Petiole 4–16 mm long, 2–5 mm wide. Leaf lamina 160–450 mm long, 43–118 mm wide, length:width ratio (2.1) 2.8–3.9, Base acute, apex acute to acuminate, surface below glabrate when young, glabrous when old; midrib sparsely pubescent to glabrous below; secondary veins 15–24 pairs. Flower pedicel 0–5 mm long. Bracts 1 to 6, upper bract 10–25 mm long, 14–25 mm wide. Sepals 11–23 mm long, 13–26 mm wide. Outer petals 20–39 mm long, 17–26 mm wide. Inner petals 20–42 mm long, 15–26 mm wide, length:width ratio 1.3–2.1, broadly obovate to obovate. Carpels 20 to 70. Fruiting pedicel ca. 5 mm long. Monocarps (only unripe fruits seen).

Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Cameroon, Equatorial Guinea (Bioko Island), Nigeria.
Habitat and ecology
Submontane to mountain mature or old secondary rain forests. Altitude: 800–1400 m a.s.l.
Additional specimens examined
Cameroon – South-West Region • A. Dahl 622 (K), trail north of Likombe village; 4°14'N, 9°11'E; alt. 1060 m; 02 Mar. 1995 • B.-A. Nkongmeneck 891 (YA), Mt Cameroun, flanc d'Ekona Lelu, feuille IGN: 1/200 000 Buea-Douala; 4°16'N, 9°18'E; alt. 1300 m; 14 Jan. 1985 • D.W. Thomas 4469 (K, MO, P, P, YA), forest and meadows on the gently sloping side of Mt Cameroun above small Koto village; 4°18'N, 9°06'E; alt. 550 m; 06 Mar. 1985 • E.W.G. Kalbreyer 41 (K); 4°01'N, 9°12'E; 1877 • G.W.J. Mildbraed 10720 (K), Likomba – Pflangzung, 15–35 km NE of Victoria; 4°06'N, 9°20'E; alt. 50 m; Nov. 1928 • H. Lehmbach 178 (B), Buea; 4°09'N, 9°14'E; alt. 1800 m; 16 Jan. 1898 • H. Lehmbach 57 (B, K), Buea1, Buea; 4°09'N, 9°14'E; alt. 1000 m; 26 Apr. 1897 • J.J. Wieringa 2058 (WAG); Fako, Mt Etinde (=Small Mt Cameroon), near and at summit; 4°05'N, 9°07'E; alt. 1400 m; 29 Jan. 1994 • M. Groves 122 (K, SCA, YA), trail north of Likombe Village; 4°14'N, 9°11'E; alt. 1000 m; 21 Feb. 1995 • M.R. Cheek 5145 (K, SCA, YA), Liwenyi, on the West bank of the Onge river above the first set of rapids, that is about 1–2 hours walk inland from Enyenge; 4°17'N, 8°58'E; alt. 50 m; 27 Oct. 1993 • P. Lane 142 (K, SCA, YA), Mt Kupé, within Permanent Sample Plot on Shrike Trail leading from Nyasoso to summit; 4°50'N, 9°40'E; alt. 1200 m; 20 Jun. 1994 • R.W.J. Keay FHI37485 (FHI, K), Kumba Distr., eastern boundary of Bambuko F.R., about 11 miles SSW. of Musome. Northern slopes of Cameroon Mt; 4°18'18.11'N, 9°11'55.75'E; alt. 1000 m; 01 Feb. 1958 • S. Cable 1353 (K, SCA, YA), path towards grassland and top of Mt Cameroon from Likombe; 4°07'N, 9°11'E; alt. 1060 m; 22 Feb. 1995 • S. Cable 1524 (K, SCA, YA), Upper Boando, logging path that becomes trail to summit; 4°04'N, 9°09'E; alt. 900 m; 14 Mar. 1995 • S. Cable 1626 (K, YA), path to summit of Etinde from Upper Boando; 4°04'N, 9°09'E; alt. 1000 m; 16 Mar. 1995 • S. Cable 2187 (K, YA); Ndian, Korup National Park, Ekundu Kundu, path from Ekundu-Kundu to about 1 km; 5°02'14.24'N, 8°54'58.14'E; alt. 300 m; 26 Apr. 1996 • T.D. Maitland 453 (K), Buea, above upper farm; 4°09'N, 9°14'E; alt. 1219 m; 1924 • T.D. Maitland s.n.10 (K), Cameroon mountain, Buea; 4°09'N, 9°14'E; alt. 975 m; 1930 • T.D. Maitland s.n.9 (K), Cameroon mountain, Buea area; 4°09'N, 9°14'E; alt. 1219 m; 1930 • T.L.P. Couvreur 1026 (WAG, YA), on trail leading to top of Mt Etinde, after Ekonjo village; 4°04'02.27'N, 9°09'10.16'E; alt. 749 m; 01 Apr. 2016 • T.L.P. Couvreur 1029 (MPU, WAG, YA), on trail leading to top of Mt Etinde, after Ekonjo village; 4°04'04.83'N, 9°09'11.43'E; alt. 781 m; 01 Apr. 2016 • T.L.P. Couvreur 1040 (MPU, WAG, YA), Mt Cameroon National Park, Bakinguili trail, above Bakinguili village; 4°05'48.05'N, 9°03'24.71'E; alt. 563 m; 02 Apr. 2016 • T.L.P. Couvreur 1046 (MPU, WAG, YA), Mt Cameroon National Park, on the Bomona trail, behind Bomona village, 10 km NW from Idenau; 4°17'46.46'N, 9°06'06.31'E; alt. 859 m; 03 Apr. 2016 • T.L.P. Couvreur 990 (MPU, WAG, YA), slopes of Mt Cameroon, on the Bokwango trail, near Bokwango village, 4 km south west of Buea; 4°07'25.26'N, 9°11'11.28'E; alt. 1227 m; 23 Mar. 2016 • T.L.P. Couvreur 992 (MPU, WAG, YA), slopes of Mt Cameroon, on the Bokwango trail, near Bokwango village, 4 km south west of Buea; 4°07'27.06'N, 9°10'13.93'E; alt. 1560 m; 23 Mar. 2016. Equatorial Guinea – Bioco (Fernando Poo) • G.W.J. Mildbraed 6428 (B), Pico Basilé, Fernando Poo: Nordseite d. Pics v. Sta. Isabel oberhalb Basilé, Wald über der Kakao-Region 6–800 m, viel Allanblackia, (oberer Tropenwald); 3°41'24.72'N, 8°51'27.72'E; alt. 800 m; 16 Aug. 1911 – Bioko Sur • W.R.Q. Luke 12203 (MA), Moeri: Camp1 to Camp2; 3°28'09.84'N, 8°40'06.96'E; alt. 704 m • W.R.Q. Luke 13279 (K), Hormiga Camp pt 346 to North camp pt 347; 3°20'51'N, 8°29'23.64'E; alt. 800 m; 01 Feb. 2009. Nigeria – Cross River State • K. Schmitt 323 (MO), Cross River National Park. Oban Hills. SW facing slope ca 5 km E of Neghe; 5°15'20“N, 8°38'50"E; 07 Feb. 1995.
Uvariodendron fuscum var. giganteum
≡ Uvaria gigantea
Engl., Notizbl. Königl. Bot. Gart. Berlin 2: 292. 1899 (quoad specimens Zenker 108 and 698); Uva gigantea Kuntze, Deutsche Bot. Monatsschr. xxi. 173 (1903); Uvariodendron giganteum (Engl.) R.E.Fr., Acta Horti Berg. 10: 62 (1930). Type. Cameroon – Central Region • G.A. Zenker 108 (lectotype: P! (P00362654), designated by
Description
Young branches with long soft hairs producing a whitish appearance quickly falling off. Petiole 8–35 mm long, 3.5–8 mm wide. Leaf lamina 357–676 mm long, 84–225 mm wide, length:width ratio 2.5–4, base acute to rounded, apex acute to acuminate, surface below pilose to glabrous when young, glabrous when old; midrib pilose to glabrous below; secondary veins 22–33 pairs. Flower pedicel 0–7.5 mm long. Bracts 1 to 6, upper bract 16–22 mm long, 20–50 mm wide. Sepals 20–30 mm long, 16–26 mm wide. Outer petals 25–40 mm long, 17–30 mm wide. Inner petals 21–40 mm long, 17–29 mm wide, length:width ratio 1.1–1.5, broadly obovate to obovate. Carpels 50 to 100. Fruiting pedicel 9–15 mm long. Monocarps 6 to 24, 20–50 mm long, 11–25 mm wide, length:width ratio 1.7–3.8, cylindrical, curved, acuminate, slightly constricted between the seeds, puberulent, green to brown. Seeds 8–14 per monocarp, ca. 14 mm long, ca. 7 mm wide.
Distribution
Element of the Lower Guinean Domain and Congolia Domain of the Guineo-Congolian Region: Cameroon, Democratic Republic of the Congo, Gabon.
Habitat and ecology
Lowland to submontane mature or old secondary rain forests, on inundated soils or along streams or rivers. Altitude: 100–1300 m a.s.l.
Notes
Ud. fuscum var. giganteum differs from the type variety in having young branches and petiole covered with long soft hairs producing a whitish appearance quickly falling off (vs. young branches and petiole sparsely pubescent to glabrous). Compared to the type variety, it has larger leaves (35.7–67.6 cm long and 8.4–22.5 cm wide, vs. 16–45 cm long and 4.3–11.8 cm wide), with generally more secondary veins (22 to 33 vs. 15 to 24), and flowers with generally greater sepals (20–30 mm long vs. 11–23 mm long) and more carpels (50 to 100 vs. 20 to 70) (Fig.
Additional specimens examined
Cameroon – Central Region • G.A. Zenker 698 (B), Yaoundé, Yaunde; 3°52'N, 11°31'E; alt. 800 m; 04 Feb. 1895 • T.L.P. Couvreur 419 (MPU, WAG, YA), Mont Mbam Minkon, on trail, 5 km from Nkol Nyada village; 3°58'04.91'N, 11°24'08.64'E; alt. 1000 m; 21 Mar. 2013 – East Region • T.L.P. Couvreur 1206 (MPU, WAG, YA), 60 km south of Yokadouma, 30 km after Ngato, 15 km after river. ALPICAM ‘base de vie', then on forestry road starting 4 km before Maséa village; 3°09'55.18'N, 14°42'25.64'E; alt. 587 m; 05 Mar. 2019 • T.L.P. Couvreur 1229 (MPU, WAG, YA), 60 km south of Yokadouma, 30 km after Ngato, 15 km after river. ALPICAM ‘base de vie', then 40 km on forestry road starting 4 km before Maséa village. Coupe 4–4 of UFA 023; 3°05'08.17'N, 14°40'18.2'E; alt. 622 m; 08 Mar. 2019 – South Region • G.A. Zenker 1438 (L), Bipinde, Urwaldgebiet; 3°05'N, 10°25'E; 1898 • M. Biholong 279 (P, YA), près d'Alati. Ancienne piste Alati – Mintom II; 2°11'44.45'N, 13°24'17.61'E; 17 Jan. 1975 – South-West Region • C. Doumenge 473 (L, MO, P), Cameroon. Bakossi Mountains 1–8 Km NNE of Menyum Village; 5°01'N, 9°38'E; alt. 1000 m; 22 May. 1987 • P. Lane 501 (K, SCA, WAG, YA), Mt Kupe Division. Ndum. Forest trail 2 km south from Etube-Tape village; 4°51'N, 9°42'E; alt. 1200 m; 02 Feb. 1995 • T.L.P. Couvreur 1057 (MPU, WAG, YA), Nyasoso village, on max's trail to Mt Kupe; 4°49'27.88'N, 9°41'59.42'E; alt. 1227 m; 05 Apr. 2016 • T.L.P. Couvreur 512 (MPU, YA); Fako, on trail from Ekongo village, located 5 km before the entrance to Limbe, 7 km on secondary road. On flank of Mt Etinde. 100 m in Mont Cameroon National Park; 4°04'19.23'N, 9°08'00.76'E; alt. 959 m; 16 Oct. 2013 • T.R. van Andel 3761 (MO, SCA, U, WAG), Bokwango. Trail to Mann's spring; 4°07'44'N, 9°11'26'E; alt. 1130 m; 21 Jun. 2001. Democratic Republic of the Congo – Orientale • T.B. Hart 910 (WAG); Mambasa, Epulu, Zaire, Zone de Mambasa (Ituri); 1°25'N, 28°35'E; alt. 750 m; 23 Mar. 1989. Gabon – Estuaire • T.-J. Klaine 1690 (P), Environs de Libreville; 0°25'N, 9°27'E; 18 Oct. 1899 – Ogooué-Ivindo • N. Hallé 3156 (P), Bélinga; 1°05'N, 13°08'E; 12 Nov. 1964 • N. Hallé 430 (K, P), Bélinga, mines de fer, le long de la rivière Folley; 1°07'N, 13°11'E; 12 Aug. 1966 • N. Hallé 549 (K, P), Bélinga, mines de fer; 1°08'N, 13°12'E; alt. 700 m; 16 Aug. 1966.
Uvariodendron fuscum var. magnificum , stat. nov.
≡ Uvariodendron magnificum Verdc. syn. nov., Kew Bull. 23(3): 515 (1969). Type. Uganda – Western Province • Okodi in Hamilton 696 (holotype: K! (K000198899, K000198900, K000198896, K000198897, K000198898); isotypes: ENT, MHU! (MHU000022, MHU000023, MHU000024)); Ankole District, Kashoya • Kitomi CFR; 0°13'N, 30°15'E; alt. 1050 m; 15 Jun. 1968.
Description
Young branches with long soft hairs producing a whitish appearance quickly falling off. Petiole 6–20 mm long, ca. 5 mm wide. Leaf lamina 210–750 mm long, 57–215 (250) mm wide, length:width ratio 3–4, base acute to cuneate, apex rounded to acuminate, surface below pilose to glabrous when young, glabrous when old; midrib pilose to glabrous below; secondary veins 23–30 pairs. Flower pedicel 10–15 mm long. Flower not seen, description from
Distribution
Endemic to Zambezian Region. Known from only one locality in the Western Province in Uganda.
Habitat and ecology
Montane forest. Altitude: around 1050 m a.s.l.
Notes
Ud. fuscum var. magnificum differs from the type variety in having young branches and petioles covered with long soft white hairs producing a whitish appearance quickly falling off (vs. young branches and petiole sparsely pubescent to glabrous). Compared to the type variety, it has larger leaves (21.–75 cm long and 5.7–21.5 (25) cm wide, vs. 16–45 cm long and 4.3–11.8 cm wide), with more secondary veins (23 to 30 vs. 15 to 24). It differs from the type variety and from the var. giganteum by its flowers having longer pedicels 10–15 mm long (vs. 0–7.5 mm long), larger sepals (30–55 mm long and 30–43 mm wide, vs. 11–30 mm long and 13–26 mm wide), larger petals (50–70 mm long and 36–47 mm wide, vs. 20–40 mm long and 15–30 mm wide) and more carpels (150 to 160 vs. 20 to 100) (Fig.
Additional specimens examined
Uganda – Western Province • T. Synnott 197 (MHU); Ankole District, North border of Kitomi Forest; 0°07'29.07'N, 30°18'07.11'E; alt. 1070 m; 22 Oct. 1968.
Uvariodendron gorgonis
Type
Kenya – Coast • B. Verdcourt 3940 (holotype: K! (K000198893), sheet here designated; isotype: BR! (BR0000008824325), EA! (EA000002458, EA000002460, EA000002459), K! (K000198894, K000198892, K000198895); Kwale District, Mrima hill (about halfway up road to Lungalunga from Mwambweni); 4°29'06.39'S, 39°15'47.19'E; alt. 182 m; 16 Jan. 1964.
Description
Tree 3–27 m tall, D.B.H. 3–50 cm; young branches pubescent to glabrous, old branches glabrous; plant with lemon smell. Leaf bud ‘eragrostiform’, composed of 6–12, distichous, longitudinally folded, velutinous scales. Petiole 4–18 mm long, 1.5–5 mm wide, sparsely pubescent to glabrous. Leaf lamina 153–410 mm long, 36–127 mm wide, length:width ratio 2.3–4.3, elliptic to oblong to obovate, coriaceous, base generally acute to rounded, and minutely decurrent at the extreme base, apex generally acute to acuminate but can also appear rounded or retuse, acumen 0.5–12 mm long; surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 13–22 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and branches, composed of 1–3 flowers. Flower pedicel 8–18 mm long, 3–8 mm in diameter, velutinous. Flowers bisexual, buds globose, sessile, 7–15 mm high, 9–14 mm in diameter, velutinous. Bracts 1 to 8, upper bract 3–12 mm long, 9–25 mm wide, broadly ovate, clasping the pedicel, enclosing the bud, velutinous outside, glabrous inside. Sepals 3, 8–13 mm long, 8–24 mm wide, imbricate to fused over up to ca. 30 % of their length, velutinous outside, glabrous inside, clear brown. Outer petals 3, 15–35 mm long, 13–23 mm wide, length:width ratio 1.1–2.2, broadly elliptic to elliptic, velutinous outside, glabrous inside, cream outside, cream with a dark purplish red streak from base up to 75% of the petal length inside. Inner petals 3, 15–40 mm long, 9–16 mm wide, length:width ratio 1.5–2.7, obovate, valvate at apex, velutinous outside, glabrous inside, cream to cream with a dark purplish red streak at base up to 15% of their length outside, dark purplish red with cream apex inside. Stamens 1400 to 1600, 2.5–3 mm long, 0.4–1 mm wide, anthers linear, connective prolongation truncate. Carpels 40 to 80, 3–4.5 mm long, 0.8–1.5 mm wide, pubescent, free; stigma 0.5 mm long, 0.8–1.1 mm wide, coiled, velutinous. Fruiting pedicel 10–20 mm long, 3–5 mm in diameter, velutinous. Monocarps 20 to 60, 24–90 mm long, 4.5–11 mm wide, length:width ratio 5–11, very narrowly cylindrical, torulose to torose, velutinous, greyish green; stipe 5–11 mm long, 1.5–3 mm wide, velutinous. Seeds 3–14 per monocarp, uniseriate, 3 to 9 mm long, 2–7 mm wide, ellipsoid.

Uvariodendron gorgonis Verdc A habit B trunk and branches, note the young fruits all along the branches and trunk C young branches and leaves, upper side D young fruit, side view E flower, bottom view F flower, side view, inset: Coleoptera larvae frequently found in the flower. A, B no specimen associated C Dagallier 52 D, E Dagallier 38. Photos Léo-Paul Dagallier.
Distribution
Endemic to Somalia-Masai Region: Kenya, Mozambique and Tanzania.
Habitat and ecology
Lowland to submontane secondary rain forests or semi-deciduous forest. Soil: coral rag or igneous intrusion. Altitude: 170–950 m a.s.l.
Phenology
Flowers collected from January to February and from July to November. Fruits collected from January to March and from June to November.
Vernacular names
Tanzania: ‘Mtwamu’ (Mbago 1616).
Etymology
Although

Uvariodendron gorgonis Verdc A young branch and apical ‘eragrostiform’ bud B flower, top view, and flower buds, side view C longitudinal section of flower bud D stamen, side view E carpel, side view F longitudinal section of carpel G fruit, side view H seed, side view. A from Verdcourt 1890 B–G from Drummond 1954 H from Verdcourt 3940 (type). Drawings by Heather Wood, from
Notes
This species differs from all other species by its fruits, unique in the genus. The fruits of Ud. gorgonis are composed of 20 to 60 monocarps, while most of the Uvariodendron species have fruits composed of 1 to 20 monocarps (except Ud. calophyllum and Ud. fuscum var. giganteum that have fruits composed of up to 35 monocarps). Moreover, the monocarps are 5 to 10 times longer than wide (vs. 1.3 to 4.5 times longer than wide in the other species). They are torulose to torose, i.e. very strongly constricted between the seeds. The ovules (and seeds) are uniseriate. The combination of these characters confers the fruits the aspect of a hairy head.

Vegetatively, this species is harder to differentiate as its leaves have dimensions that overlap with most of the other species. However, the bases of the leaves are quite peculiar: they appear generally acute to rounded but a closer look actually detects them as minutely decurrent at the extreme base. Similarly, the apices of the leaves are generally acute to acuminate, but can appear rounded to slightly retuse with a minute acumen.
The fresh leaves of Ud. gorgonis also present a citrus smell when crushed, as in Ud. angustifolium, Ud. citriodorum and Uvariopsis citrata.
Preliminary conservation status
A previous assessment, that needs updating, listed this species as Endangered EN under criteria B2ab(iii), based on its occurrence in eight locations in Tanzania (Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project 2009a). Here, the EOO of this species is estimated at 203,044 km2 and its AOO at 72 km2. It occurs in 11 locations in Tanzania, Kenya and Mozambique. Given its habitat is severely fragmented and there is continuing decline due to habitat loss and degradation (Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project 2009a), it qualifies for the same previous status of Endangered EN under criteria B2ab(iii).
Additional specimens examined
Kenya – Coast • B. Verdcourt 1890 (K); Kwale District, Mrima Forest, Mrima Hill; 4°29'06.39'S, 39°15'47.19'E; 06 Sep. 1957 • B. Verdcourt 1911 (B, EA, K); Kwale District, Mrima Forest, Mrima Hill; 4°29'06.39'S, 39°15'47.19'E; 07 Sep. 1951 • J.P.M. Brenan 14601 (EA, K, P); Kilifi District, K7, Kilifi District. Pangani ‘Kaya' Forest; 3°51'S, 39°40'30'E; alt. 170 m; 19 Nov. 1978 • R.B. Faden 70/253 (K); Kwale District, Mrima Hill, about halfway betwee Mgambweni and Lungalunga; 4°29'S, 39°16'E; alt. 70 m; 25 Jun. 1970 • S.A. Robertson MDE78 (K); Kwale District, Mrima Hill; 4°29'S, 39°16'E; alt. 270 m; 04 Feb. 1989. Mozambique – Cabo Delgado • W.R.Q. Luke 9983 (MO), Quirimba NP, Ngura Inselberg; 12°51'S, 39°39'E; alt. 420 m; 10 Dec. 2003. Tanzania – Lindi • F.M. Mbago 2148 (K), Ruawa Forest; 9°44'46.39'S, 39°33'04.77'E; alt. 250 m; 01 Sep. 2001 – Morogoro • A. Pócs 6061B (DSM); Morogoro Rural District, Kimboza Forest Reserve, between Mkuyuni and Matombo; 7°00'S, 37°48'E; alt. 300 m; 19 Nov. 1969 • A. Pócs 6280F (DSM); Morogoro Rural District, Kimboza Forest Reserve; 7°00'S, 37°48'E; alt. 300 m; 05 Nov. 1970 • B.J. Harris 3223 (DSM, EA); Morogoro Rural District, Kimboza Forest Reserve, KFR near Matombo, Uluguru Mts; 7°00'S, 37°48'E; 06 Sep. 1969 • C.D. Mgaza 323 (K), Luzunguru forest reserve; 6°05'S, 37°40'E; 19 Sep. 1959 • F.C. Magogo 2171 (K); Morogoro Rural District, Kimboza Forest Reserve, Kibungo Forest Reserve (Kimboza); 7°00'S, 37°48'E; alt. 600 m; 31 Aug. 1981 • F.M. Mbago 1616 (DSM); Kilombero District, Lower Kihansi Hydropower Project; 8°24'S, 36°21'E; 30 Aug. 1997 • F.M. Mbago 1734 (K), T7: Udzungwa scarp, Upper Kihansi Gorge, forest opposite the spray zone; 8°34'54.35'S, 35°51'05.9'E; alt. 950 m; 15 Jul. 1998 • L.-P.M.J. Dagallier 47 (DSM, MPU, P, WAG); Morogoro Rural District, Kimboza forest; 7°01'16.37'S, 37°48'17.38'E; alt. 278 m; 15 Nov. 2019 • L.-P.M.J. Dagallier 52 (DSM, K, MPU, P, WAG); Morogoro Rural District, Kimboza forest; 7°01'18.37'S, 37°48'31.63'E; alt. 262 m; 15 Nov. 2019 • L.-P.M.J. Dagallier 57 (DSM, MPU, P, WAG); Mvomero District, Turiani village; 6°09'36.37'S, 37°36'17.72'E; alt. 377 m; 18 Nov. 2019 • L.-P.M.J. Dagallier 61 (DSM, MPU, P, WAG); Mvomero District, Turiani village; 6°09'46.37'S, 37°36'12.66'E; alt. 373 m; 18 Nov. 2019 • L.B. Mwasumbi 19144 (MO); Kilombero District, Kihansi gorge; 8°35'S, 35°51'50'E; alt. 800 m; 04 Sep. 1998 • R.B. Drummond 1954 (K); Morogoro Rural District, 6.4 km N of Turiani, Lusunguru Forest Reserve, near Mtibwa sawmill; 6°05'S, 37°41'E; 31 Mar. 1953 • S. Paulo 163 (EA, K); Morogoro Rural District, Kimboza Forest Reserve; 7°00'S, 37°48'E; Nov. 1953 • S.R. Semsei 1498 (K), Mtibwa Forest Reserve; 6°08'56.37'S, 37°50'10.32'E; Nov. 1953 • S.R. Semsei 773 (K); Morogoro Rural District, Kimboza Forest Reserve; 7°00'S, 37°48'E; Jul. 1952 • S.R. Semsei 852 (K); Morogoro Rural District, Mtibwa Forest Reserve; 6°07'S, 37°39'E; Aug. 1952 • T.L.P. Couvreur 69 (DSM, MO, WAG); Kilombero District, Kimboza Forest reserve, 2 km after Kimboza village, 45 km from Morogoro; 7°01'19.37'S, 37°48'16.8'E; alt. 250 m; 25 Nov. 2006 • Y.S. Abeid 2651 (EA, MO); Mvomero District, Kanga Forest Reserve, near Kwa Beku subvillage; 5°57'15'S, 37°43'11.4'E; alt. 500 m; 11 Mar. 2006 – Pwani • L.B. Mwasumbi 113377 (DSM); Kisarawe District, Pugu hills Forest Reserve; 6°54'S, 39°05'E; 28 May. 2002 – Tanga • F.M. Mbago 3756 (DSM); Handeni District, T3: Handeni Kwansisi Mgulwi stream; 5°51'55.38'S, 38°35'39.34'E; alt. 203 m; 16 Aug. 2016 • G.A. Peter 13872 (B, K, WAG); Lushoto District, O. Usambara, Mashewa; 4°47'S, 38°17'E; alt. 400 m; Sep. 1915 • L.-P.M.J. Dagallier 38 (BR, DSM, K, MO, MPU, P, WAG); Handeni District, Kwamsisi village; 5°51'50.38'S, 38°35'31.77'E; alt. 196 m; 13 Nov. 2019.
Uvariodendron kimbozaense , sp. nov.
Type
Tanzania – Morogoro • L.-P.M.J. Dagallier 49 (holotype: MPU! (MPU1379108); isotypes: DSM!, MPU! (MPU1375359), P! (P00948153); also distributed to K and WAG); Morogoro Rural District, Kimboza forest; 7°01'16.37'S, 37°48'35.24'E; alt. 279 m; 15 Nov. 2019.
Diagnosis
Uvariodendron kimbozaense resembles Ud. kirkii by the elliptic leaves and petal colors, but is distinguished from this species by having slightly greater leaves (140–220 mm, vs. 70–190 mm long in Ud. kirkii) with a base acute to rounded (vs. acute to decurrent). Compared to Ud. kirkii, Ud. kimbozaense also has more bracts on the flower pedicel (1 to 4, vs. 1 maximum), greater (6–12 mm long, vs. 3–6.5 mm long) and imbricate (vs. connivent) sepals, and greater (16–39 mm long, vs. 10–20 mm long), elliptic (vs. ovate), and almost flat petals (with a slight transversal curvature vs. “boat-shaped” petals, with a strong transversal curvature) (Fig.
Description
Tree 5–7 m tall, D.B.H. 15–20 cm; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 4–5 mm long, 1.5–2 mm wide, glabrous. Leaf lamina 147–215 mm long, 44–68 mm wide, length:width ratio 2.5–3.6, elliptic to oblong, coriaceous, base acute to rounded, apex attenuate, surface above glabrous, surface below glabrous; midrib slightly raised with a central groove all along above, raised below, glabrous above, glabrous below; secondary veins 12–17 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on growth of the trunk and on old branches, composed of 1–11 flowers. Flower pedicel 10–14 mm long, 2–3 mm in diameter, pubescent. Flowers bisexual, buds globose to oblate, pedicellate, 5–10 mm high, 7–16 mm in diameter, pubescent. Bracts 1 at base and from 1 to 4 along the pedicel, upper bract 3–6 mm long, 6–14 mm wide, broadly ovate, adpressed, clasping the pedicel, pubescent outside, glabrous inside. Sepals 3, 6–12 mm long, 12–21 mm wide, depressed ovate, imbricate, pubescent outside, glabrous inside, green to brownish-green. Outer petals 3, 16–28 mm long, 13–19 mm wide, length:width ratio 1.2–1.8, broadly elliptic to elliptic, puberulent outside, puberulent at apex to glabrous inside, cream outside, cream with a purple-red to reddish black streak from base up to 50% of the petal length inside. Inner petals 3, 18–39 mm long, 9–18 mm wide, length:width ratio 2–2.2, obovate, slightly transversally curved, puberulent outside, glabrous inside, cream to cream with a slight purplish red streak at base outside, purple-red at base to reddish black toward the apex inside. Stamens 600 to 800, 2–3.5 mm long, 0.5–0.7 mm wide, anthers linear, connective prolongation truncate. Carpels 11 to 16, 4–5 mm long, 1.8–2 mm wide, densely pubescent, free; stigma 1–1.5 mm long, 1.5–2 mm wide, coiled, densely pubescent, covered with an exudate at anthesis. Fruits unknown.

Uvariodendron kimbozaense Dagallier & Couvreur A habit B trunk with borne inflorescences C young inflorescence with flower buds D flower with one outer petal and the 3 inner petals torn or gnawed E young branch with leaves, lower side F young branch with leaves, upper side G detail of flower receptacle with stamens and stigmas, note the gleaming exudate on the stigmas and the hairs stuck on them H base of leaf, upper side, note the slightly raised midrib. A–H Dagallier 49 (type). Photos Léo-Paul Dagallier.
Distribution
Endemic to Somalia-Masai Region. Known from only one locality in Tanzania: the Kimboza Forest Reserve.
Habitat and ecology
Lowland mature rain forest on coral rag and limestone rocks. Altitude: 250–450 m a.s.l.
Phenology
Flowers collected in March and November.
Etymology
The specific epithet comes from Kimboza Forest Reserve (Tanzania), from where the species is endemic.
Notes
Ud. kimbozaense has been previously identified as Ud. kirkii in all the specimens examined. However, it clearly differs from Ud. kirkii by the characters presented in the diagnosis above (see Fig.
Preliminary conservation status
This species is only known from the Kimboza Forest Reserve in Tanzania. This protected area has been threatened by encroachment, logging and invasion by the exotic Cedrela odorata L. (
Additional specimens examined
Tanzania – Morogoro • C.J. Kayombo 5357 (MO); Morogoro Rural District, Kimboza Forest Reserve; 7°00'43'S, 37°48'50'E; alt. 400 m; 21 Mar. 2006 • T.L.P. Couvreur 71 (DSM, MO, WAG); Kilombero District, Kimboza Forest reserve, 2 km after Kimboza village, 45 km from Morogoro; 7°01'19.37'S, 37°48'16.8'E; alt. 250 m; 25 Nov. 2006 • W.R.Q. Luke 766 (EA), Uluguru Mts nr Ruvu bridge; 7°01'S, 37°49'E; alt. 450 m; 04 Nov. 1987.

Uvariodendron kirkii
Type
Kenya – Coast • B. Verdcourt 3939 (holotype: K! (K000198893), sheet here designated; isotypes: BR! (BR0000008824325), EA!, K! (K000198894, K000198895)); Kwale District, Mrima hill (about halfway up road to Lungalunga from Mwambweni); 4°29'06.39'S, 39°15'47.19'E; alt. 182 m; 16 Jan. 1964.
Description
Shrub to tree 1.5–6 (10–15) m tall, D.B.H. 3–20 cm; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 2.5–8 mm long, 1–2 mm wide, glabrous. Leaf lamina 70–188 mm long, 22–75 mm wide, length:width ratio 2.1–3.5, elliptic, coriaceous, base acute to decurrent, apex attenuate, surface above glabrous, surface below glabrous; midrib slightly impressed above, raised below, glabrous above, glabrous below; secondary veins 8–16 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences mostly axillary or borne on old branches, sometimes borne on trunk, composed of 1–2 flowers. Flower pedicel 5–28 mm long, 1–4 mm in diameter, pubescent. Flowers bisexual, buds globose, pedicellate, 2–5.5 mm high, 1.3–6 mm in diameter, pubescent. Bracts 1 at base and sometimes 1 on the upper half of the pedicel, upper bract 1–5 mm long, 1–8 mm wide, broadly ovate, adpressed, semi-clasping the pedicel, pubescent outside, glabrous inside. Sepals 3, 3–6.5 mm long, 4–8 mm wide, very broadly to broadly ovate, connivent, pubescent outside, glabrous inside, green to brownish-green. Outer petals 3, 10–17 mm long, 6–13 mm wide, length:width ratio 1.2–1.7, broadly ovate to ovate, pubescent to puberulent outside, puberulent to glabrous inside, cream to greenish pale yellow outside, cream to cream with a purple streak from base up to 50% of the petal length inside. Inner petals 3, 12–20 mm long, 7–8 mm wide, length:width ratio 1.3–1.9, obovate, strongly transversally curved (“boat-shape”), valvate at apex, puberulent outside, glabrous inside, white to cream outside, cream to cream with purple streak at base inside. Stamens 1000 to 1200, 1.5–1.8 mm long, 0.2–0.3 mm wide, anthers linear, connective prolongation truncate. Carpels 8 to 16 (20), 2–3 mm long, 1–1.5 mm wide, pubescent, free; stigma 0–1.5 mm long, 1–1.3 mm wide, coiled, pubescent. Fruiting pedicel 7–22 mm long, 2–4 mm in diameter, pubescent. Monocarps 1 to 6, 23–36 mm long, 10–16 mm wide, length:width ratio 1.4–2.4, cylindrical, longitudinally ridged, slightly constricted between the seeds, minutely acuminate, puberulent, green to dull orange to greyish dark blue; sessile to shortly stipitate, stipe 0–3 mm long, 2–5 mm wide, pubescent. Seeds 12–15 per monocarp, biseriate, 11 to 15 mm long, 8–10 mm wide.
Distribution
Endemic to Somalia-Masai Region: Kenya and Tanzania.
Habitat and ecology
Lowland mature or secondary rain forest, dry forest or thicket on coral rag and limestone rocks, or igneous intrusions. Altitude: 10–80 m a.s.l.
Phenology
Flowers and fruits collected all year.
Vernacular names
Tanzania: ‘Kisambaa’ in Mkomboa (Mwangoka 1356), ‘Mdumi’ in Zigua (Tanner 3467).
Notes
This species is quite variable morphologically with leaves from 70 mm to almost 200 mm long. It can be distinguished from the other species with its pedicelate flowers growing mainly on leafy branches and sometimes on the trunk. The pedicel is lower than in Ud. anisatum (5–28 mm vs. 15–65 mm long in Ud. anisatum). At anthesis, the flowers generally have their outer petals completely spread out and their inner petals connivent at apex (Fig.

Uvariodendron kirkii Verdc A flowering branch B flower, top view C stamen D longitudinal section of carpel E carpel, semi-side view F carpel, top view G fruit, side view H seed, side view I flower, side view J flower, top view K old fruit with A single rotten monocarp. A from Verdcourt 3939 (type) B–F from Drummond 4029 G, H from Drummond 3983 I, J Dagallier 23 K Dagallier 60. Drawings by Heather Wood, from
Preliminary conservation status
A previous assessment, which needs updating, listed this species as Vulnerable VU under criteria B1ab(iii) (Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project 2009b). Here, the EOO of this species is estimated at 67,203 km2 and its AOO at 152 km2. It occurs in more than 10 locations in Tanzania and Kenya and is fairly common but its population is severely fragmented and there is continuing decline due to habitat loss (Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project 2009a). We thus assign a preliminary updated conservation status of Endangered EN B2ab(iii).
Additional specimens examined
Kenya – Coast • B. Verdcourt 1891 (K); Kwale District, Mrima Forest, Mrima Hill; 4°29'06.39'S, 39°15'47.19'E; 06 Sep. 1957 • B. Verdcourt 5280 (K); Kwale District, Jombo Mt (near bottom); 4°26'08.39'S, 39°12'42.61'E; 11 Apr. 1978 • F.C. Magogo 768 (K); Kwale District, Shimba Hills, Dzombo Mountain; 4°26'09.39'S, 39°12'46.1'E; alt. 305 m; 08 Apr. 1968 • I.R. Dale 3836 (K), Witu Forest Reserve, Gongoni Forest, Witu; 2°23'07'S, 40°30'09'E; Oct. 1937 • J.B. Gillett 19901 (K); Kwale District, Diani forest. Areas NW & NE within 1 km N of turn off from new road for Jadini hotel, W & E of this road. Areas SW & SE within 1.5 km s of this point; 4°19'S, 39°33'E; alt. 12 m; 11 Jul. 1972 • J.B. Gillett 19906 (EA, K); Kwale District, Diani forest. Areas NW & NE within 1 km N of turn off from new road for Jadini hotel, W & E of this road. Areas SW & SE within 1.5 km s of this point; 4°19'S, 39°33'E; alt. 12 m; 11 Jul. 1972 • J.B. Gillett 24009 (B, EA); Kwale District, Kenya: K7: Kwale District Waa Kaya forest near coast N. of Tiwi ruined Mosque; 4°12'S, 39°36'E; alt. 30 m; 30 Dec. 1982 • J.P.M. Brenan 14592 (K); Kilifi District, K7, Kilifi District. Pangani ‘Kaya' Forest; 3°51'S, 39°40'50'E; alt. 170 m; 19 Nov. 1978 • L.J. Lap 112 (MO, WAG); Kilifi District, Kaya Kauma. Mapsheet K198/1. Along the path extending westwards from the Kaya cemetery. Western hill slope forest of Kaya Kauma; 3°37'S, 39°44'E; 22 Dec. 1981 • M.G. Gilbert 4967 (EA, K, WAG), Kwale District. Jombo Hill; 4°28'S, 39°12'E; alt. 300 m; 07 Jan. 1978 • M.G. Gilbert 5334 (K); Kilifi District, Cha Simba rocks on Mariakani – Kilifi Road; 3°44'S, 39°42'E; alt. 200 m; 16 Feb. 1979 • N. Mwadime 204 (EA); Kwale District, Shimoni, mwamba-wanga area; 4°38'30.39'S, 39°24'12.97'E; alt. 2 m; 13 Dec. 2012 • R.B. Drummond 3983 (K); Kwale District, Jardini Beach, 18 miles south of Mombasa, sea level; 4°19'30'S, 39°34'20'E; 26 Aug. 1953 • R.B. Drummond 4029 (K, U); Kwale District, Mwasangombe Forest, 15 miles S.W. of Kwale; alt. 230 m; 27 Aug. 1953 • R.B. Faden 77/397 (K); Kwale District, Kinondo (Ngalani Kaya) Forest; 4°23'30'S, 39°33'30'E; alt. 5 m; 15 Feb. 1977 • R.B. Faden 77/437 (K); Kilifi District, just north of Mwarakaya on Chonyi – Ribe road; 3°47'S, 39°42'E; alt. 140 m; 16 Feb. 1977 • S. Rawlins EAH11275 (K), Witu; 2°23'05.40'S, 40°30'04.68'E; alt. 24 m; Jun. 1957 • S.A. Robertson 5929 (K); Kwale District, Kaya Diani; 4°16'S, 39°35'E; alt. 10 m; 10 Nov. 1989 • S.A. Robertson 6465 (EA); Kilifi District, Watamu, Ashe plot; 3°20'S, 40°01'E; alt. 10 m; 28 May. 1991 • S.A. Robertson 7550 (EA, K, WAG); Kilifi District, Kambe Rocks; 3°51'S, 39°40'E; alt. 120 m; 13 May. 2005 • S.A. Robertson MDE293 (EA, K, WAG), Kwale Distr., Dzombo Hill; 4°26'S, 39°13'E; alt. 400 m; 09 Feb. 1989 • S.A. Robertson MDE46 (K); Kwale District, Mrima hill; 4°29'S, 39°16'E; 03 Feb. 1989. Tanzania – Dar es Salaam • J. Kirk s.n (K), Dar es Salaam; 6°48'57.39'S, 39°16'49.44'E; Mar. 1860 – Kaskazini Pemba (North Pemba) • P.J. Greenway 2681 (K); Micheweni District, Ras Kigomasha; 4°52'S, 39°41'E; 10 Dec. 1930 – Lindi • K.B. Vollesen 3150 (EA), c. 7 km NWN of Kingupina; 8°24'S, 38°31'E; alt. 125 m; 25 Dec. 1975 • R.I. Ludanga 1369 (EA), Selous, Likandage (Utunge); 9°00'S, 37°30'E; 28 Jan. 1972 – Morogoro • L.-P.M.J. Dagallier 60 (DSM, MPU, P, WAG); Mvomero District, Turiani village; 6°09'40.37'S, 37°36'14.48'E; alt. 384 m; 18 Nov. 2019 – Pwani • B.J. Harris 6209 (EA), Approx 6 mls north of Chalinze on road D'Salaam-Chalinze-Segera; 6°33'23.38'S, 38°19'08.14'E; alt. 274 m; 04 Apr. 1972 • G.A. Peter 24366 (B, K), Useguha; Aug. 1918 – Tanga • G.A. Peter 40249 (B), Usaguha: Hale in Pangani; 5°18'S, 38°36'E; alt. 330 m; 14 May. 1926 • G.A. Peter 52280 (B); Usambara, E Usambara; 5°00'45.38'S, 38°40'43.59'E; 05 Aug. 1915 • H.G. Faulkner 11 (B, K); Tanga District, Sawa Creek; 5°07'S, 39°06'E; 17 Feb. 1957 • J. Procter 3592 (K); Pangani District, Msumbugwe Forest Reserve; 5°30'15.38'S, 38°45'23.76'E; Apr. 1967 • L.-P.M.J. Dagallier 23 (DSM, MPU, P, WAG), Kilulu Hill, 50 km North of Tanga; 4°46'21.39'S, 39°07'33.48'E; alt. 247 m; 08 Nov. 2019 • L.-P.M.J. Dagallier 8 (DSM, K, MPU, P, WAG), forest patch near Msangazi river, along the road from Chalinze to Korogwe-Tanga; 5°34'16.38'S, 38°26'47.01'E; alt. 306 m; 06 Nov. 2019 • L.-P.M.J. Dagallier 9 (DSM, MPU, P, WAG), Kilulu Hill, 50 km North of Tanga; 4°46'23.39'S, 39°07'29.07'E; alt. 263 m; 07 Nov. 2019 • M.A. Mwangoka 1356 (MO); Muheza District, Kuze Kibago village. NW part of Kwemnyese public forest patch; 4°54'51'S, 38°43'35'E; alt. 100 m; 01 Jun. 2000 • R.E.S. Tanner 12 (K); Pangani District, Msubugwe Forest; 5°31'59.38'S, 38°43'59.88'E; alt. 100 m; 1955 • R.E.S. Tanner 29 (K); Pangani District, Msubugwe Forest; 5°31'59.38'S, 38°43'59.88'E; alt. 100 m; 1955 • R.E.S. Tanner 3467 (B, K); Pangani District, Mwera, Kwa Besa, Mwanamgaru; 5°29'S, 38°57'E; 28 Mar. 1957 • R.E.S. Tanner 6 (K); Pangani District, Msubugwe Forest; 5°31'59.38'S, 38°43'59.88'E; alt. 100 m; 1955 • T.L.P. Couvreur 34 (DSM, FHO, MO, NHT, WAG); Handeni District, road 3 km before arriving to Kwedikwazu coming from Dar Es Salaam. At Msangasi river; 5°34'21.38'S, 38°26'46.8'E; alt. 300 m; 14 Nov. 2006 • W.D. Hawthorne 776 (K), Nr Langoni village; 5°28'S, 38°54'E; 27 May. 1982 – Zanzibar Central/South • J. Vaughan 2318 (BM), Kufile cave; 6°25'08.39'S, 39°29'15.85'E; 23 Feb. 1936 – Zanzibar West • D. Aplin 1 (K), Zanzibar: Chumbe I; 6°16'44'S, 39°10'37'E; alt. 5 m; Feb. 1988 • J. Vaughan 1689 (K), Kombeni caves; 6°15'39.39'S, 39°15'39.95'E; 22 Nov. 1930 • J. Vaughan 1715 (K), Kombeni caves; 6°15'39.39'S, 39°15'39.95'E; 04 Dec. 1930 • J. Vaughan 1731 (EA, K), Kombeni caves; 6°15'39.2'S, 39°15'39.95"E; Dec. 1930 • J.H. Vaughan 1689 (K); Magharibi (West) District, Kombeni Cave Well; 6°15'S, 39°16'E; 22 Nov. 1930 • P.J. Greenway 2654 (K); Magharibi (West) District, Haitajwa Hill; 6°16'S, 39°16'E; 04 Dec. 1930 – Unknown major area • Botany Students DSM1394 (DSM, K, WAG), Steinbruch Forest Reserve near Maweni, W of Tanga; 5°06'S, 39°01'E; alt. 80 m; 31 Dec. 1969 • G.A. Peter 4487 (B, K, WAG), Useguha; 6°00'S, 38°15'E; 10 Jun. 1914.

Uvariodendron mbagoi
Type
Tanzania – Tanga • L.-P.M.J. Dagallier 39 (holotype: MPU! (MPU1375316); isotypes: DSM!, MPU! (MPU1375317), P! (P00948145, P00948146); also distributed to K, MO, WAG); Handeni District, Kwedijela forest, ~8 km Kwamsisi village; 5°54'50.38'S, 38°36'12.35'E; alt. 156 m; 13 Nov. 2019.
Description
Tall shrubs to trees 3–6 m tall, D.B.H. 5–10 cm; young branches sparsely pubescent to glabrous, old branches glabrous; slash with strong bergamot smell (the citrusy smell of Citrus bergamia Risso). Leaves stiff, greyish green, with margin slightly revolute. Petiole 3–6.5 mm long, 1.2–3 mm wide, sparsely pubescent to glabrous. Leaf lamina 76–157 mm long, 31–59 mm wide, length:width ratio 2.2–3.5, narrowly elliptic to elliptic to obovate, between coriaceous and cartilaginous, base acute to slightly decurrent (sometimes cuneate), apex acute to shortly acuminate, acumen 5–10 mm long; surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous when young, glabrous when old below; secondary veins 10–13 pairs, weakly brochidodromous, indistinct to slightly impressed above, (slightly) raised below; tertiary veins reticulate. Inflorescences borne on trunk and old branches, composed of 1–2 (3) sessile flowers. Flower pedicel 0–0.6 mm long, ca 2 mm in diameter, velutinous. Flowers bisexual, buds globose, 5–9 mm high, 5–9 mm in diameter, velutinous, falling off very easily. Only flower buds and old fallen flowers seen. Bracts 2 to 5 at base of the pedicel, upper bract 4–8 mm long, 10–15 mm wide, appressed, enclosing the bud, pubescent outside, glabrous inside. Sepals 3, ca. 7–8 mm long, ca. 7–12 mm wide (measures taken from bud), imbricate, enclosing the petals in bud, velutinous outside, glabrous inside, brown. Outer petals 3, 4.5–5.5 mm long, 4.5–5 mm wide (measures taken from bud), length:width ratio ca. 1, ovate, shortly velutinous outside, glabrous inside, color unknown. Inner petals 3, 3.5–4.5 mm long, 4–4.5 mm wide (measures taken from bud), length:width ratio ca. 1, ovate, glabrous outside, glabrous inside, color unknown. Stamens 400 to 500, mature length unknown, anthers linear, connective prolongation truncate. Carpels 12 to 16, ca. 1.5 mm long, ca. 1 mm wide (measures taken from old flower), velutinous, free; stigma mature length unknown, coiled. Fruiting pedicel 0–6 mm long, ca. 4 mm in diameter, pubescent. Monocarps 1 to 7, 20–50 mm long, 10–12 mm wide, length:width ratio 2–4.5, cylindrical, generally curved, longitudinally ridged and slightly constricted between the seeds, tomentose with regular tufts of high hair density, green-grey; sessile to shortly stipitate, stipe 0–1.5 mm long, ca. 5 mm wide, tomentose. Seeds 4–17 per monocarp, uniseriate to biseriate, 8 to 8.5 mm long, 5.5–6 mm wide, ellipsoid.

Uvariodendron mbagoi Dagallier & Couvreur A trunk with old or fallen flowers B young branch with leaves, upper side C leaf, lower side D leaf, upper side E bract remaining from A fallen flower (top) and flower bud (bottom) F detail of A moncarp, note the hair pattern G fruit, semi-top view H tangential (left) and longitudinal (right) cuts of A monocarp. A, C Dagallier 40 B, D–H Dagallier 39 (type). Photos Léo-Paul Dagallier.
Distribution
Endemic to Somalia-Masai Region: Tanzania.
Habitat and ecology
Lowland rain forest or dry forest. Soil: coral rag. Altitude: 90–340 m a.s.l.
Phenology
Flowers collected from August to December. Fruits collected from August to November.
Vernacular names
Tanzania: ‘Mkenene’ in Chizigua (Bloesch s.n.; Couvreur 3; Dagallier 39, 40).
Uses
The bark is used as spice for tea or meat meals (Dagallier 39).
Notes
This species differs from the other Uvariodendron species by the strong bergamot scent of crushed fresh and dry leaves and bark, by having stiff greyish-green leaves with slightly revolute margins, and by its globose flower buds easily falling off and its tomentose fruits having regular tufts of higher hair density.

Preliminary conservation status
Following IUCN criterion B, this species has been previously assigned a preliminary status of EN B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv) (
Additional specimens examined
Tanzania – Morogoro • L.-P.M.J. Dagallier 50 (DSM, K, MPU, P, WAG); Morogoro Rural District, Kimboza forest; 7°01'18.38“S, 37°48'32.13"E; alt. 267 m; 15 Nov. 2019 – Pwani • L.-P.M.J. Dagallier 1 (DSM, K, MPU, P, WAG), Msata Hill, 30 km North of Chalinze; 6°22'17.78“S, 38°21'49.97"E; alt. 317 m; 06 Nov. 2019 • L.-P.M.J. Dagallier 3 (WAG), Msata Hill, 30 km North of Chalinze; 6°22'17.09“S, 38°21'50.23"E; alt. 328 m; 06 Nov. 2019 • L.-P.M.J. Dagallier 6 (WAG), Msata Hill, 30 km North of Chalinze; 6°22'17.93“S, 38°21'49.94"E; alt. 323 m; 06 Nov. 2019 • T.L.P. Couvreur 3 (DSM, WAG); Bagamoyo District, Mazizi hill, on road between Chilinze and Wami River; 6°22'14.4“S, 38°21'51"E; alt. 100 m; 09 Nov. 2006 • U. Bloesch s.n (WAG), Kwedijela Coastal Forest, T3; 5°55'S, 38°36'E; 18 Sep. 2004 – Tanga • C.M. Kisena 3039 (MO); Handeni District, T3; Collected from Kwedivikilo sacred forest near Manga Village; 5°06'S, 38°37'E; 17 Nov. 1997 • F.M. Mbago 3323 (DSM, K); Handeni District, Kwedijela forest, 8 km Kwamsisi village; 5°51'S, 38°33'E; 07 Oct. 2004 • G.A. Peter 52283 (B, K, WAG); Pangani District, Useguha. Inseln des Pangani bei Hale; 5°17'34.8“S, 38°36'14.06"E; alt. 340 m; 31 Jan. 1915 • L.-P.M.J. Dagallier 40 (DSM, K, MO, MPU, P, WAG); Handeni District, Kwedijela forest, ~8 km Kwamsisi village; 5°54'50.77“S, 38°36'13.27"E; alt. 155 m; 13 Nov. 2019 • T.C.E. Congdon 532 (K); Pangani District, Mkwaja Ranch; 5°48'50.76“S, 38°47'40.92"E; alt. 90 m; 04 Dec. 1998 • W.D. Hawthorne 1420A (K), Mkulumuzi river, karst river valley, Steinbruch reserve; 5°06'S, 39°01'00.12»E; 12 Aug. 1982.
Uvariodendron molundense
≡ Uvaria molundensis Diels, Bot. Jahrb. Syst. 53(3–5): 435 (1915). Type. Cameroon – East Region • G.W.J. Mildbraed 4373 (holotype: B! (B 10 0153118)), Südkameruner Waldgebiet: Bezirk Molundu, ‘Bange Busch’, unbewohnter Urwald zwischen Lokomo, Bumba und Bange; 2°50'N, 15°15'E; 29 Jan. 1911.
= Uvaria letestui
Pellegr., Bull. Mus. Natl. Hist. Nat. xxvi. 658 (1920); Uvariodendron letestui (Pellegr.) R.E.Fr., Acta Horti Berg. 10: 60 (1930). Type. Gabon – Nyanga • G.M.P.C. Le Testu 1234 (holotype: P! (P00315833), sheet designated by
= Uvaria mayumbensis Exell, J. Bot. 64(Suppl. 1): 3 (1926); Uvariodendron mayumbense (Exell) R.E.Fr., Acta Horti Berg. 10: 57 (1930). Type. Angola – Cabinda • J. Gossweiler 6159 (holotype: BM! (BM000554073)), Mayombe Pango Mariga [Maringa?]; 4°55'S, 12°25'E; 17 Jan. 1916.
= Uvaria mannii Hutch. & Dalziel, Fl. W. Trop. Afr. I. 50 (1927); Kew Bull 150 (1927). Type. Equatorial Guinea – Bioko Norte • G. Mann 257 (holotype: K! (K000198802); isotype: P! (P00315831)); 3°40'N, 8°47'E; 1860.
Description
Tree to shrub 1.2–10 m tall, D.B.H. 1.5–20 cm; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 4–17 mm long, 1.5–5 mm wide, sparsely pubescent to glabrous. Leaf lamina (180) 200–460 mm long, 51–160 mm wide, length:width ratio 2.2–4.8, elliptic to oblong to obovate, coriaceous, base acute to rounded (sometimes minutely decurrent at the very base), apex attenuate to acuminate, acumen 2–38 mm long; surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 12–22 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and branches or axillary, composed of 1 flower. Flower pedicel 2–14 mm long, 1–4 mm in diameter, pubescent. Flowers bisexual, buds globose, sessile, 3.5–9 mm high, 4–14 mm in diameter, velutinous. Bracts 1 to 6, upper bract 3–8 mm long, 3–10 mm wide, broadly ovate, clasping the pedicel, velutinous outside, glabrous inside. Sepals 3, 5–9 (14) mm long, 5–10 mm wide, imbricate (sometime fused at base), velutinous outside, glabrous inside, green to brown. Outer petals 3, 12–30 mm long, 9–22.5 mm wide, length:width ratio 1.1–2.5, elliptic, pubescent outside, pubescent at apex to glabrous inside, light yellow to cream outside, purple with cream margins inside. Inner petals 3, 11–27 mm long, 8–20 mm wide, length:width ratio 1.2–2.4, obovate, pubescent outside, glabrous inside, light yellow to cream outside, purple with cream margins inside. Stamens 1000 to 1200, 0.8–2.5 mm long, 0.1–1 mm wide, anthers linear, connective prolongation truncate. Carpels 8 to 40, 1–5 mm long, 0.5–2 mm wide, pubescent, free; stigma 0–2 mm long, 1–1.5 mm wide, coiled, densely pubescent, covered with an exudate at anthesis. Fruiting pedicel 3–15 mm long, 2.5–6 mm in diameter, pubescent to glabrous. Monocarps 2 to 10, 21–60 mm long, 9–30 mm wide, length:width ratio 1.3–3.4, cylindrical, generally straight, truncate or rounded at apex, pubescent to glabrate, greyish green to orange; sessile to shortly stipitate, stipe 0–4 mm long, 2–4 mm wide, pubescent to glabrate. Seeds 3–10 per monocarp, biseriate, 12 to 14 mm long, 7–10 mm wide, semicircular, pale brown to pinkish brown.

Uvariodendron molundense (Diels) R.E.Fr. A leaves, lower side (left) and upper side (right), note the morphological variation B leaf bases, upper side, note the morphological variation C young branch with leaves D stamen, front view E detail of flower showing pedicel bracts and sepals, bottom view, not the imbrication of sepals F flower, top view G detail of flower showing stamens and stigmas, note the gleaming exudate on the stigmas H flower buds, side view I young fruit with unripe monocarps, top view J ripe monocarp, side view K monocarp, transversally open L branches with borne flowers. A (left) E–H Bidault 2222 A (right) Couvreur 655 B (top) Couvreur 542 B (bottom) C Bidault 4269 D Couvreur 1195 I Bidault 2248 J, K Couvreur 932 L Couvreur 1172. Photos A (left) B (bottom) C, E–H, I Ehoarn Bidault A (right) B (top) J–L Thomas Couvreur D Léo-Paul Dagallier.
Distribution
Element of the Lower Guinean Domain and Congolia Domain of the Guineo-Congolian Region: Cameroon, Democratic Republic of the Congo, Gabon, Equatorial Guinea (Bioko Island), Republic of the Congo.

Uvariodendron molundense (Diels) R.E.Fr. A leaf B flower on branch, side view C flower, bottom view D flower, petals open, top view E longitudinal section of receptacle F stamen, front view G carpel, side view and detail of ovules H floral diagram I fruit, longitudinal sections of monocarps A–C, E–G from Le Testu 9649 D from Le Testu 8437 H, I from Hallé 3264. Drawings by Hélène Lamourdedieu, from
Habitat and ecology
Lowland and premontane mature and old secondary rain forests. Altitude: 0–1000 m a.s.l.
Phenology
Flowers and fruits collected all year.
Notes
This species presents a great morphological variability in terms of leaf size and shape. Vegetatively it resembles Ud. connivens and Ud. fuscum (especially var. fuscum), although it generally has smaller leaves than these two species. In flower, it differs in having a short pedicel 2–14 mm long (vs. a long pedicel 10–40 mm long in Ud. connivens), imbricate sepals (vs. fused at base in Ud. fuscum), smaller petals (11–30 mm long vs. 20–70 in Ud. fuscum), and fewer carpels (eight to 40 vs. 20 to 160 in Ud. fuscum).

Conservation status
This species has been assessed as Least Concern LC (Botanic Gardens Conservation International and IUCN SSC Global Tree Specialist Group 2019a).
Additional specimens examined
Cameroon – Central Region • D. Dang 643 (P, YA), colline ‘ZOABISSIM' dans l'apellation locale, 4 km envirion N du village EKEKAM; 3°55'N, 11°22'E; alt. 1085 m; 09 Mar. 1978 • F. Tadjouteu 566 (K, YA), Ndanan 1, Mefou National park, 18 km from MFOU; 3°33'N, 11°36'02.47'E; alt. 700 m; 22 Mar. 2004 • M.R. Cheek 11839 (YA), Ndanan 1, Eastern part of the park, West of Ndangan 1; 3°37'19'N, 11°36'03'E; alt. 710 m; 18 Mar. 2004 – East Region • J.F. Villiers 625 (P, YA), Camp CFA Bango I, bord de la riv. Bango 36 km SE Bateka Malen, village situé à 21 km N de Molundou (Feuille IGN Molundou 1/200.000); 2°17'49.56'N, 15°09'41.81'E; 08 Nov. 1971 • R.G. Letouzey 3117 (P, YA), Bertoua, Kap; 4°35'N, 13°40'50'E; 23 Feb. 1960 • R.G. Letouzey 5142 (P, YA), a 30 km au NE de Bangé (km75 route Yokadouma – Molundu) feuille IGN 1/200 000. Yokadouma; 3°00'25.8'N, 15°08'13.99'E; 25 May. 1963 • T.L.P. Couvreur 1195 (MPU, WAG, YA), 60 km south of Yokadouma, 30 km after Ngato, 15 km after river. ALPICAM ‘base de vie', then on forestry road starting 4 km before Maséa village; 3°09'19'N, 14°44'16.87'E; alt. 613 m; 04 Mar. 2019 – Littoral • T.L.P. Couvreur 1172 (MPU, P, WAG, YA), Mapubi, 30 km before Edea on Yaoundé-Edea road. On forestry road, 5 km direction to Sanaga river; 3°50'39.26'N, 10°23'22.88'E; alt. 164 m; 28 Feb. 2018 • T.L.P. Couvreur 613 (MPU, YA), Ebo Wildlife Reserve, Djuma permanent camp. On Djashaka trail; 4°21'06.18'N, 10°14'12.1'E; alt. 378 m; 13 Feb. 2014 • T.L.P. Couvreur 652 (WAG, YA), Mambe Massif, above Boga village, 100 km along road from Yaoundé to Edea; 3°54'36.26'N, 10°46'31.86'E; alt. 637 m; 19 Jun. 2014 • T.L.P. Couvreur 655 (MPU, WAG, YA), Mambe Massif, above Boga village, 100 km along road from Yaoundé to Edea; 3°54'29.55'N, 10°46'20.29'E; alt. 657 m; 19 Jun. 2014 • T.L.P. Couvreur 656 (WAG, YA), Mambe Massif, above Boga village, 100 km along road from Yaoundé to Edea; 3°54'24.79'N, 10°46'17.82'E; alt. 653 m; 20 Jun. 2014 • W.J.J.O. de Wilde 1420 (K, MO, P, WAG), 40 km NW. of Eséka, W. of Yaoundé; 3°51'N, 10°32'E; 12 Dec. 1963 – South Region • B.-A. Nkongmeneck 800 (YA); Océan, environs de Mvini, à 34 km E de Campo. Feuille IGN 1/200 000 Kribi-Nyabessa; 2°22'18.12'N, 10°06'15.48'E; 24 Oct. 1984 • G.A. Zenker 3511 (BM, K, L, P), Kamerun, Bipinde; 3°05'N, 10°25'E; 1908 • G.P. Tchouto Mbatchou T8X70 (WAG), Campo Ma'an area, Bibabimvoto, Dipikar island, in the Campo area along transect T8; 2°16'16'N, 10°03'35'E; alt. 60 m; 26 Aug. 2000 • G.W.J. Mildbraed 5936 (B, K), S. Cameroon: Kribi; 2°48'N, 10°24'E; 1911 • J.J. Bos 3259 (P, WAG, YA), About 13 km from Kribi, Ebolowa road; 2°51'N, 10°00'E; 13 Nov. 1968 • J.J. Bos 5474 (BR, MO, P, WAG, YA), S. bank of Lobé river, SE. of Gr. Batanga ferry; 2°52'N, 9°54'E; 11 Oct. 1969 • J.J. Bos 6521 (WAG), 6 km N. of km 46 Kribi – Lolodorf; 3°05'N, 10°15'E; 12 Mar. 1970 • J.J. Bos 7075 (P, WAG), 5 km N. of km 7 Kribi – Ebolowa road, behind Pygmee village; 2°56'N, 9°57'E; 10 Jul. 1970 • M.E. Elad 1270 (KRIBI, WAG), Campo-Ma'an area, Bibabimvoto, along transect T4; 2°15'03'N, 10°21'51'E; alt. 40 m; 01 Feb. 2000 • T.R. van Andel 4228 (KRIBI, U, WAG, YA), Campo Ma'an area, Boussebeliga. Trail to Bimvoa chute; 2°43'N, 9°52'E; alt. 40 m; 26 Oct. 2001 – South-West Region • B. Sonké 1226 (K, SCA); Ndian, Dikome; 4°27'N, 9°01'E; 05 May. 1994 • B.N. Khayota 537 (K, SCA, YA), Upper Boando, footpath to south of village; 4°05'N, 9°10'E; alt. 650 m; 14 Mar. 1995 • D. Kenfack 1509 (MO), Mokoko, at the center base forest; 4°27'N, 9°04'00.12'E; 25 Apr. 2001 • G.P. Tchouto Mbatchou 136 (K, SCA, YA), Mapanja; 4°05'N, 9°09'E; alt. 1040 m; 20 Apr. 1992 • G.P. Tchouto Mbatchou 611 (K, SCA, YA), Mokoko, forest above Bonja village; 4°28'N, 9°06'E; alt. 240 m; 23 Mar. 1993 • M. Akogo 234 (K, SCA, YA), Ekumbe Mofako, Mokoko Forest Reserve, Ekumbe-Mofako; 4°28'N, 9°03'E; alt. 140 m; 21 Apr. 1994. Central African Republic – Sangha-Mbaéré • D.J. Harris 5112 (E, MO), Kongana research camp, 25 km SE of Bayanga; 2°47'N, 16°25'E; 08 Jun. 1994 • D.J. Harris 5504 (E, MO), Kongana camp, 22 km SE of Bayanga; 2°47'N, 16°26'E; 11 Feb. 1996. Democratic Republic of the Congo – Bas-Congo • P. Compère 1112 (K), vallée de la Quinongo. Territ: Songololo. Province: Léopoldville; 5°09'57.14'S, 14°19'37.82'E; 21 Dec. 1959 – Orientale • P. Gérard 5604 (K, WAG); Ango, Digba, Foret de Akare entre rivière Bili et Ase; 4°23'N, 25°48'E; 05 Nov. 1963. Equatorial Guinea – Bioko Sur • W.R.Q. Luke 11889 (EA), Moaba. Pt 140; 3°14'06'N, 8°37'22'E; alt. 8 m; 15 Mar. 2007. Gabon – Haut-Ogooué • G.M.P.C. Le Testu 9649 (P); 1°38'S, 13°35'E; 30 Oct. 1930 – Moyen-Ogooué • T.L.P. Couvreur 928 (LBV, WAG, YA), 27 km after Lambaréné, on road to Bifoum (N1), then around 20 km on road to Lake Azingo; 0°27'52.7'N, 10°02'18.45'E; alt. 95 m; 24 Nov. 2015 • T.L.P. Couvreur 932 (LBV, WAG, YA), 33 km south of Lambaréné, then 18 km on dirt road just after Mimongo village, in Morel petrol consession; 1°02'10.10'S, 10°23'39.95'E; alt. 185 m; 25 Nov. 2015 – Ngounié • G.M.P.C. Le Testu 5762 (BM, P), Mbigou, Mboumi riverside; 1°55'S, 11°55'E; 18 Nov. 1925 • G.V. Dauby 574 (LBV, MO), Est du Parc National de Waka, à ± 5 km au Sud de la rivière Mayi; 1°13'36.11'S, 11°17'09.31'E; alt. 604 m; 17 Feb. 2008 • J.C. Arends 450 (LBV, WAG), Waka River; 1°13'S, 10°52'E; alt. 330 m; 25 Nov. 1984 • O.L.S. Lachenaud 1444 (BRLU, LBV, MO), Mabounié, piste du nord-est; 0°43'02'N, 10°36'05'E; 17 Nov. 2013 • P.J.M. Maas 10207 (LBV, WAG), side roads of road Pény-Mouila, in CBG concession; 2°00'45.10'S, 10°28'52.7'E; alt. 115 m; 10 Nov. 2011 – Nyanga • E. Bidault 4269 (BR, BRLU, LBV, MO, P, WAG), Entre Ndendé et le Congo, au Sud de Nzinga, bords de la rivière Douvono; 3°04'56.11'S, 11°39'34.31'E; alt. 154 m; 04 Apr. 2018 • G.M.P.C. Le Testu 1211 (BM), Tchibanga area, Nganda; 2°50'S, 11°00'E; 31 Oct. 1907 – Ogooué-Ivindo • A. Moungazi 1631 (LBV, MO, P, WAG), Ivindo National Park, route Langoué, rivière Niadou; 0°15'N, 12°25'E; 05 May. 2004 • A. Moungazi 223 (P), Makokou, 10 km S de Makoukou; 0°30'30.82'N, 12°47'45.59'E; alt. 500 m • E. Bidault 2222 (BRLU, LBV, MO), concession forestière Rougier-Ivindo; 0°05'34'N, 12°21'15'E; alt. 283 m; 27 Oct. 2015 • E. Bidault 2248 (BR, BRLU, LBV, MO, MO, P, WAG), concession forestière Rougier-Ivindo; 0°04'05'N, 12°21'15'E; alt. 460 m; 28 Oct. 2015 • J. Florence 930 (P), Station d'Ipassa, 10 Km S de Makokou; 0°30'N, 12°47'E; alt. 500 m; 14 Apr. 1978 • N. Hallé 3264 (L, P, U), Bélinga; 1°05'N, 13°08'E; 19 Nov. 1964 • N. Hallé 3332 (P), Bélinga; 1°05'N, 13°08'E; 25 Nov. 1964 • N. Hallé 431 (MO, P, WAG), Bélinga, mines de fer, le long de la rivière Folley; 1°07'N, 13°11'E; 12 Aug. 1966 • N. Hallé 451 (MO, P), Bélinga, mines de fer, route du Belvédère; 1°05'N, 13°12'E; alt. 900 m; 12 Aug. 1966 • T.L.P. Couvreur 564 (YA), along main trail departing from behind the herbarium at the Research station of Ipassa, in Ivindo National Park; 0°30'13.77'N, 12°47'36.4'E; alt. 519 m; 11 Nov. 2013 – Ogooué-Lolo • A.M. Louis 648 (WAG), ± 70 km N.W. of Lastoursville, along rd. to Achouka, old forest along Lolo R. near Wagny settlement; 0°36'N, 12°19'E; alt. 250 m; 13 Nov. 1983 • F.J. Breteler 13331 (WAG), near Bambidie, E of Lastoursville; 0°44'N, 12°58'E; alt. 300 m; 16 Oct. 1994 • G.M.P.C. Le Testu 49 (P), region de Lastoursville, Lastoursville; 0°50'N, 12°42'E; 1934 • G.M.P.C. Le Testu 8437 (BM, P), region de Lastoursville, Nzocou; 1°28'S, 12°28'E; 12 Oct. 1930 • J.J. Wieringa 6223 (LBV, MO, WAG), c.55 km N of Lastoursville, CEB forestry concession ‘Milolé' (UFA2-UFG2-lot2), foothills of Ngota Mountain; 0°20'24'N, 12°46'48'E; alt. 330 m; 28 Jan. 2008 • J.J. Wieringa 6263 (LBV, MO, WAG), c.30 km ENE of Lastoursville, 15 km on forestry road from Bambidie to Akieni; 0°39'11.4'N, 12°56'36.6'E; alt. 320 m; 29 Jan. 2008 • T.L.P. Couvreur 1110 (LBV, WAG), Lastoursville, in Precious Woods conssesion (CEB). 30 km east of Bambidie village, 55 km from Lastoursville; 0°39'46.04'N, 12°45'55.58'E; 07 Jun. 2016 – Ogooué-Maritime • M.S.M. Sosef 2270 (LBV, WAG), Doudou Mountains National Parc, c. 5 km S of Camp Peny (CBG); 2°03'30'S, 10°28'12'E; alt. 100 m; 14 Nov. 2005 • T.L.P. Couvreur 539 (MPU, YA), 5 km on loggers road from Peny (CGB) village; 2°01'55.10'S, 10°29'59.48'E; alt. 188 m; 07 Nov. 2013 • T.L.P. Couvreur 542 (MPU, YA), 5 km on loggers road from Peny (CGB) village; 2°01'53.10'S, 10°29'45.29'E; alt. 200 m; 07 Nov. 2013 – Woleu-Ntem • G.M.P.C. Le Testu 9064 (P), region de Minvoul, Minvoul; 2°10'N, 12°10'E; Mar. 1933 • L. Ngok Banak 1570 (LBV, LBV, MO, P, WAG), Minkébé National Park, southern inselberg area; 1°23'10.2'N, 12°33'17.4'E; alt. 620 m; 02 May. 2003 • M.S.M. Sosef 1894 (WAG), forestry concession Bordamur, c. 40 km NE of Mitzic; 1°00'06'N, 11°51'12'E; alt. 500 m; 06 Feb. 2003 • MINKébé Series AM60 (WAG), Minkébé area, at 2000 m from the camp; 1°30'N, 12°48'E; 25 Mar. 1990 • MINKébé Series V150 (WAG), Minkébé area, 10 x 10 m inventory plot V, placed 3.5–13.5 m north at 200–210 m on transect A; 1°30'N, 12°48'E; 04 Mar. 1990 • MINKébé Series W15 (WAG), Minkébé area, near base camp number 1; 1°29'N, 12°49'E; 08 Feb. 1990 • MINKébé Series W403 (WAG), Minkébé area, 800 m on transect B (to the south); 1°30'N, 12°48'E; 20 May. 1990 • T.L.P. Couvreur 853 (LBV, WAG, YA), Oyem, on road to Mbolonzok (off main road to Mongono and Equatorial Guinea); 1°32'39.18'N, 11°30'43.04'E; alt. 660 m; 13 Nov. 2015 • T.L.P. Couvreur 876 (LBV, WAG, YA), on road from Mitzic to Lalara (N2), just after the bridge over the Lara, c.500 m in forest; 0°36'12.83'N, 11°29'11.53'E; alt. 580 m; 15 Nov. 2015. Republic of the Congo – Kouilou • C. Farron 4882 (P), Pointe Noire, route du chantier de Boungolo (Pointe-Noire); 4°07'S, 11°57'E; 31 Jan. 1966 – Mianga N'Bissy 183 (P), Moyen Congo, Mayombe central, vers km 57 chemin de fer Pointe Noire – Brazzaville; 4°29'27'S, 12°12'15'E; Sep. 1947 • P. Sita 3759 (P), route du nouveau campement Maamar sur 18 km après l'ancien; 3°45'S, 11°15'E; 09 May. 1974 • T.L.P. Couvreur 789 (IEC, WAG), 30 km on Dolisie-Mvouti road, just behind the telephone antenna; 4°12'49.12'S, 12°31'44.71'E; alt. 710 m; 21 Sep. 2015 – Niari • P. Cabalion 153 (P), 3 km O Banga Route Vounda-Banda; 3°33'18.97'S, 11°59'56.48"E; 06 Feb. 1976 • P. Sita 3995 (P), Niari-ouest, region Banda, Vallé de la Ngouanga; 3°24'S, 12°09'E; 05 Nov. 1975.
Uvariodendron mossambicense , sp. nov.
Type
Mozambique – Manica • F. de A. Mendonça 2558A (holotype: WAG! (WAG.1418614); isotypes: COI! (COI00085396), LISC), Chimoio, Cataratas do Rio Revué; 19°37'S, 33°31'E; 23 Oct. 1944.
Diagnosis
Ud. mossambicense closely resembles Ud. dzomboense by its narrowly elliptic and small (less than 140 mm long) leaves with acute to slightly decurrent base. However, Ud. mossambicense has flower buds ca. 6 mm in diameter (vs ca. 4 mm in Ud. dzomboense), covered by 2–5 velutinous bracts ca. 4 mm long (vs. 6 sparsely pubescent bracts 5–6 mm long). Ud. mossambicense has ca. 5 carpels (vs. 50–75 in Ud. dzomboense) and monocarps with stipe ca. 12 mm long (vs. sessile monocarps) (Table
Description
Probably a small tree but height unknown, D.B.H. unknown; young branches pubescent to glabrous, old branches glabrous; leaf bud ‘eragrostiform’, composed of 7, distichous, longitudinally folded, velutinous scales. Petiole 3–5 mm long, ca. 1.5 mm wide, glabrous. Leaf lamina 80–135 mm long, 30–50 mm wide, length:width ratio ca. 3, narrowly elliptic, coriaceous, base acute to slightly decurrent, apex attenuate, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 10–16 pairs, brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on branches, composed of 1 flower. Flowers bisexual, buds globose, sessile, ca. 6 mm high, ca. 6.5 mm in diameter, velutinous. Only flower bud seen. Bracts 2 to 5, upper bract ca. 4 mm long, ca. 7 mm wide, broadly ovate, appressed, half-enclosing the flower bud, velutinous outside. Sepals 3, 4–5 mm long, ca. 6 mm wide (measures taken from bud), broadly ovate, fused at base on almost 50 % of their length, forming a 3-lobed cupule, velutinous outside, glabrous inside. Outer petals 3, ca. 5 mm long, ca. 5 mm wide (measures taken from bud), length:width ratio ovate, velutinous outside. Inner petals 3, length, shape, indumentum and color unknown. Stamens numerous, length unknown, anthers linear. Carpels ca. 5, glabrous, free; stigma unknown, pubescent. Fruiting pedicel unknown. Monocarp 1 seen, ca. 37 mm long, ca. 9 mm wide, length:width ratio ca. 4.1, cylindrical, curved, slightly acuminate, glabrous, (monocarp found in the pocket of the specimen); stipitate, stipe ca. 12 mm long, ca. 3 mm wide, glabrate. Seeds 4 per monocarp, uniseriate, size and shape unknown.

Distribution
Endemic to Somalia-Masai Region. Only known from one locality in Mozambique: Chimoio, near the falls of Revué river.
Habitat and ecology
Unknown. Altitude around 100 m a.s.l.
Phenology
Flowers and fruits collected in October.
Etymology
The specific epithet comes from the country where the single specimen of this species was found.
Preliminary conservation status
This species is known from a single location, not situated in a protected area. Its EOO and AOO are thus estimated lower than 4 km2. Although the botanical exploration of the country is continuously updated (

Uvariodendron occidentale
Type
Ivory Coast – Oumé • A. Aubréville 4140 (holotype: P! (P00315840), sheet here designated; isotype P! (P00315843)), Oume, région d'Oumé; 6°23'N, 5°25'W; 28 Feb. 1957.
Description
Tree 5–10 m tall, D.B.H. up to 15 cm; young branches glabrous, old branches glabrous. Petiole 5–10 mm long, 1–3 mm wide, sparsely pubescent to glabrous. Leaf lamina 170–350 mm long, 55–95 mm wide, length:width ratio 2.5–3.7, elliptic to obovate, coriaceous, base acute to decurrent, apex acuminate, acumen 7–21 mm long; surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 10–18 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk, composed of 1 flower. Flower pedicel 6–16 mm long, 1–2 mm in diameter, pubescent. Flowers bisexual, buds globose, pedicellate, 4–6 mm high, 7–8 mm in diameter, pubescent. Bracts 1 to 3, upper bract 2.5–4 mm long, 4.5–5 mm wide, ovate, semi-clasping to clasping the pedicel, pubescent outside, pubescent inside. Sepals 3, 3–5 mm long, 2.5–6 mm wide, depressed ovate to broadly triangular, fused at base in a ring, pubescent outside, glabrous inside, color unknown. Outer petals 3, 15–25 mm long, 10–17 mm wide, length:width ratio 1.4–1.6, elliptic, pubescent outside, glabrous inside, yellow with purple marks on the margins outside, yellow with purple marks inside. Inner petals 3, 10–21 mm long, 6–10 mm wide, length:width ratio obovate, pubescent outside, glabrous inside, color unknown. Stamens 600 to 800, 2–2.5 mm long, 0.4–0.5 mm wide, anthers linear, connective prolongation truncate and puberulent. Carpels 20 to 40, 3–4 mm long, 1 mm wide, pubescent, free; stigma 0.5–1 mm long, 1 mm wide, coiled, pubescent. Fruiting pedicel 15–22 mm long, 2–3.5 mm in diameter, pubescent to glabrous. Monocarps 6 to 11, 17–50 mm long, 7–20 mm wide, length:width ratio 1.6–2.7, cylindrical, curved, longitudinally ridged, acuminate, pubescent to glabrate, color unknown; stipitate, stipe 5–12 mm long, 1–2.5 mm wide, puberulent. Seeds 9–14 per monocarp, biseriate, 7 to 5 mm long, ca. 5 mm wide.

Uvariodendron occidentale Le Thomas A leaf B inflorescence C flower, bottom view D outer petals, outside (left) and inside (right) views E inner petals, outside (left) and inside (right) views F longitudinal section of the receptacle G stamen, front view H carpel and longitudinal section I fruit, bottom view J fruit, side view K flower, one outer and one inner petals removed, bottom view. A–I from Aubréville 4140 (type) J Kouamé 1487 K Aubréville 4140. Drawings by Hélène Lamourdedieu, modified from
Distribution
Element of the Upper Guinean Domain and Lower Guinean Domain of the Guineo-Congolian Region: Ghana, Ivory Coast, Nigeria.
Habitat and ecology
Lowland mature rain forest. Altitude: 155–250 m a.s.l.
Phenology
Flowers collected from October to February. Fruits collected from January to April.
Vernacular names
Ivory Coast: ‘Michiti à grandes feuilles’ (Aubréville 4140).
Etymology
A. Le Thomas initially named this species Ud. occidentalis (

Notes
This species resembles Ud. fuscum and Ud. molundense but can be differentiated in having leaf bases acute to decurrent (vs. acute to rounded), flower slender flower pedicel 6–16 mm long and 1–2 mm wide (vs. flower subsessile in Ud. fuscum and flower pedicel rather short and thick 2–14 mm long and 2–4 mm wide in Ud. molundense), and sepals 3–5 mm long fused at base forming a ring (vs. 11–55 mm long and fused in Ud. fuscum and 5–9 mm long and free in Ud. molundense).
Preliminary conservation status
A previous assessment, that needs updating, listed this species as Vulnerable VU under criteria A1c (
Additional specimens examined
Ghana – Ashanti Region • J.E. Andoh 4171 (K, P), Ofin Headwaters Reserve; 7°00'N, 1°27'W; Apr. 1936. Ivory Coast – Bouaflé • C.C.H. Jongkind 4349 (WAG), Parc National de la Marahoue. Near south border; 6°59'N, 6°10'W; alt. 250 m; 11 Feb. 1998 – Daloa • F.N. Kouamé 1443 (CSRS, G), F.C. du Haut-Sassandra, Nord. forêt dégradée, relevé FNK14; 7°01'N, 7°18'W; 04 Apr. 1995 • F.N. Kouamé 1486 (CSRS, G), F.C. du Haut-Sassandra, Centre. forêt dégradée de pente, relevé FNK14, entre layons 10 et 11; 7°00'N, 8°10'W; 13 Apr. 1995 • F.N. Kouamé 1487 (G), F.C. du Haut-Sassandra, Centre, forêt dégradée, relevé FNK14; 7°07'45.48'N, 6°59'29.4'W; 13 Apr. 1995 – Divo • L. Aké Assi 4633 (P), Divo, Foret Classee de, Forêt de Divo; 5°54'30.6'N, 5°16'41.52'W; alt. 155 m; 11 Jun. 1958 • L. Aké Assi 6629 (G), Divo, Foret Classee de, Forêt de Divo; 5°54'30.6'N, 5°16'41.52'W; alt. 155 m; Nov. 1962 – Guiglo • A. Aubréville 1224 (P), Giglo, Debokun; 6°32'37.26'N, 7°29'36.58'W; 18 Apr. 1932 • A. Bakayoko 21 (G), Guiglo, Zaipobly, forêt sur colline; 7°29'N, 5°57'W; 27 Jan. 2001 – San-Pédro • L. Aké Assi 17192 (G), route de Tabou, forêt près de Para; 4°48'N, 6°48'W; 28 Dec. 1985 – Unknown major area • F.N. Kouamé 1623 (CSRS, G), Daloa F.C. du Haut-Sassandra, Nord. forêt très dégradée sur cuirasse, relevé FNK23; 7°18'N, 7°01'W; 07 Feb. 1995 – Unknown collector 1780 (P (P01982836)), Bords de l'Agnéby, Mudjika; Feb. 1933. Nigeria – Ondo State • F. Anakwense FHI19701 (K); Akure, Akure Forest Reserve; 7°19'N, 5°02'E; 11 Dec. 1950 • K. Obeng-Darko 182 (K); Akure, Owena Forest Reserve, Owena; 7°12'N, 5°01'E; 20 Feb. 1946 • R.W.J. Keay FHI24604 (K); Akure, Akure Forest Reserve, Block 6C Akure FR; 7°19'N, 5°02'E; 28 Oct. 1948.
Uvariodendron pilosicarpum , sp. nov.
Type
Gabon – Nyanga • J.L.C.H. van Valkenburg 2540 (holotype: WAG! (WAG.1418718); isotypes: BR! (BR0000009320413), E, K, LBV, MO, P! (P01956020), WAG! (WAG.1418719, WAG.1418720)), concession SFN; 2°39'34.2'S, 10°27'03'E; alt. 200 m; 01 Nov. 2003.
Diagnosis
Ud. pilosicarpum resembles Ud. molundense and Ud. citriodorum but can be differentiated in having the longest leaves smaller than 25 cm (vs. longest leaves greater than 25 cm long), obovate leaves (vs. elliptic to oblong to obovate leaves), with base decurrent (vs. base rounded to convex) and monocarps ellipsoid, curved, acuminate and densely pubescent (vs. monocarps cylindrical, straight and pubescent to glabrate).
Description
Tree 4–8 m tall, D.B.H. ca. 8 cm; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 4–7 mm long, 1.5–2.5 mm wide, sparsely pubescent to glabrous. Leaf lamina 178–243 mm long, 60–87 mm wide, length:width ratio 2.3–3.6, obovate, coriaceous, base decurrent, apex acuminate, acumen 13–21 mm long; surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 12–19 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and branches, composed of 1 flower. Flower pedicel unknown. Flowers bisexual, buds unknown. Mature flower unknown, measures taken from fruits. Bracts 1, ca. 10 mm long, ca. 12 mm wide, broadly ovate, semi clasping the pedicel, velutinous outside, glabrous inside. Sepals 3, 9–10 mm long, 12.5–18 mm wide (measures taken from young fruits), imbricate, pubescent outside, glabrous inside, color unknown. Outer petals unknown. Inner petals unknown. Stamens unknown. Carpels ca. 30, free. Fruiting pedicel 8–13 mm long, 3.5–5.5 mm in diameter, pubescent to sparsely pubescent. Monocarps 6 to 30, 23–32 mm long, 10–15 mm wide, length:width ratio 2.2–2.3, ellipsoid, curved, acuminate at apex, densely velutinous, bright green with brown indumentum to glaucous with silvery indumentum, with resinous fragrance when cut; sessile to shortly stipitate, stipe 0–2 mm long, 2–3 mm wide, densely pubescent. Seeds unknown.

Uvariodendron pilosicarpum Dagallier & Couvreur A full specimen sheet with young branch and leaves and fruit B fruit with some monocarps transversally cut C young fruit with remaining sepals and bract. A, B van Valkenburg 2540 (type) C Sosef 2261. Photos A Naturalis (https://data.biodiversitydata.nl/naturalis/specimen/WAG.1418718) B, C Léo-Paul Dagallier.
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Gabon.
Habitat and ecology
Lowland mature or secondary rain forest. Altitude: 100–200 m a.s.l.
Phenology
Flowers and fruits collected in November.
Etymology
The specific epithet refers to the densely pubescent monocarps, characteristic of this species.
Preliminary conservation status
This species is known from only two occurrences in the Monts Doudou National Park in Gabon. The AOO is estimated at 8 km2. Based on criterion B it would qualify for Endangered EN B1(a)+2(a), but no other subcriteria (b or c) is met. The outlook for this species relies on the future of this protected area and its distribution depends on the effects of human activities or random events in an uncertain future. Following IUCN criterion D, it is thus assigned a preliminary conservation status of Vulnerable VU D2.

Additional specimen examined
Gabon – Ogooué-Maritime • M.S.M. Sosef 2261 (BR, K, LBV, MO, WAG), Doudou Mountains National Parc, c. 5 km S of Camp Peny (CBG); 2°03'30'S, 10°28'12'E; alt. 100 m; 14 Nov. 2005.
Uvariodendron pycnophyllum
≡ Uvaria pycnophylla Diels, Bot. Jahrb. Syst. 53(3–5): 434 (1915). Type. Tanzania – Tanga • Brönnle 1876 (lectotype, here designated: FHO! (00060171); sheet from B lost or destroyed); Lushoto District, Amani; 5°06'S, 38°38'E; 20 Jul. 1908.
Description
Tree 3–10 m tall, D.B.H. 20–30 cm; young branches glabrous, old branches glabrous; bark of trunk and branches reddish, external thin layers peeling off. Petiole 8–11 mm long, 1.7–3 mm wide, glabrous. Leaf lamina 158–282 mm long, 43–83 mm wide, length:width ratio 3–4.5, elliptic to oblong, coriaceous, base acute to slightly decurrent, apex attenuate to acuminate, acumen 7–13 mm long; surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 11–17 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on old branches or axillary, composed of 1 (sub)sessile flowers. Flower pedicel 0–10 mm long, ca. 4 mm in diameter, pubescent. Flowers bisexual, buds globose to ovoid, sessile, 7–10 mm high, 7.5–12 mm in diameter, pubescent. Bracts 2 to 6, up to 9 enclosing the bud, upper bract 10–13 mm long, 16–20 mm wide, depressed ovate to broadly ovate, adpressed, clasping the pedicel and sepals, pubescent outside, glabrous inside. Sepals 3, 10–12 mm long, ca 10 mm wide, fused at base, velutinous outside, glabrous inside, brown outside, yellow with red streak at base inside. Outer petals 3, 20–36 mm long, 16–19 mm wide, length:width ratio 1.1–2, elliptic to broadly obovate, free (valvate in bud), velutinous outside, glabrous inside, brownish to reddish-yellow outside, yellow with a pinkish-red streak from base up to 80% of the petal length inside. Inner petals 3, 20–31 mm long, ca. 15 mm wide, length:width ratio 1.9–2.1, obovate, valvate at apex (imbricate in bud), puberulent outside, glabrous inside, light yellow to cream, sometimes with a slight pinkish-red streak outside, pinkish-red to dark purplish-red with yellow margins inside. Stamens 1000 to 1200, ca. 3 mm long, ca. 0.4 mm wide, anthers linear, connective truncate, yellow. Carpels 30 to 40, 4.5 mm long, 1.2 mm wide, velutinous, free; stigma 1.5 mm long, 1.2 mm wide, coiled, velutinous, covered with an exudate at anthesis. Fruiting pedicel 0–5 mm long, 3.5 mm in diameter, velutinous. Monocarps 3 to 7, 19–35 mm long, 14–22 mm wide, length:width ratio ca 1.6–2.2, ovoid to cylindrical, pubescent, red with brown pubescence, only unripe fruits seen; sessile, stipe 0–1 mm long, 2.2 mm wide, puberulent. Seeds ca. 4 per monocarp, uniseriate, unknown.

Uvariodendron pycnophyllum (Diels) R.E.Fr. A trunk, note the reddish peeling bark B young branch with leaves, upper side C young branch with leaves, lower side D young flower, top view E semi-open flower, top view F fruit, side view. A Dagallier 33 B, D–F Couvreur 21 C Dagallier 37. Photos A, C Léo-Paul Dagallier B, D–F Thomas Couvreur.
Distribution
Endemic to Somalia-Masai Region: Tanzania.
Habitat and ecology
Lowland and premontane mature rain forest. Soil: rocky or sandy soil (sandstone). Altitude: 400–1025 m a.s.l.
Phenology
Flowers collected in November. Fruits collected in April, May and November.
Notes
This species can be differentiated from other species by its bark being reddish, with external thin layers peeling off. The specimen Brönnle 1876 in B, defined as the type by

Preliminary conservation status
A previous assessment, that needs updating, listed this species as Endangered EN under criteria B1ab(iii) (Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project 2009c). Here the EOO of is estimated at 9,926 km2 and AOO at 32 km2. It is known from 12 occurrences in Tanzania, mostly situated in the East Usambara Mountains. Only two of the occurrences of this species are within protected areas, namely the Mtai Forest Reserve and the Kiwengoma Forest Reserve. Most of the remaining occurrences are less than 5 km from other protected areas (e.g. Amani Forest Reserve, Nilo Forest Reserve), and it is thus possible that this species occurs in these protected areas. However, this would need to be verified, and given the uncertainty of the occurrence of this species in protected areas other than Mtai and Kiwengoma Forest Reserves, the number of locations and area of occupancy is likely to decrease in the future. Following IUCN criterion B, it is thus assigned a preliminary updated conservation status of Endangered EN 2ab(ii,iv).
Additional specimens examined
Tanzania – Lindi • C.J. Kayombo 4558 (MO); Kilwa District, T8. NE end of Mbarawala Plateau, above valley running SE. Evergreen mixed dry rocky forest. Transect #3; 9°00'47'S, 39°07'59'E; alt. 280 m; 04 Nov. 2003 – Pwani – Frontier-Tanzania Coastal Forest Res.Prog 700 (K, MO); Rufiji District, Kiwengoma Forest. Northern edge of the Matumbi Highlands; 8°21'S, 38°56'E; alt. 365 m; 31 Jan. 1990 – Tanga • D.M. Johnson 1947 (DSM); Muheza District, along road Bombani-Kisiwani, ca 3 km from Kisiwani; 5°01'S, 38°38'24'E; alt. 400 m; 05 Jun. 1996 • F.M. Mbago 3366 (DSM); Muheza District, 3 km from Kisiwani to Bombani along the road; 5°06'38.38'S, 38°40'49.08'E; alt. 150 m; 18 May. 2005 • L.-P.M.J. Dagallier 33 (BR, DSM, K, MO, MPU, P, WAG); Muheza District, along the road to Amani Nature Reserve; 5°06'04.38'S, 38°40'30.44'E; alt. 359 m; 11 Nov. 2019 • L.-P.M.J. Dagallier 37 (DSM, K, MO, MPU, P, WAG); Muheza District, along the road to Amani Nature Reserve; 5°06'07.38'S, 38°40'16.12'E; alt. 378 m; 12 Nov. 2019 • L.B. Mwasumbi 112474 (DSM); Muheza District, T3: Kwamngumi Forest Reserve; 4°57'S, 38°43'E; 12 Mar. 1999 – Langheinrich FHDS2889 (B); Korogwe District, W Usambara Mts., Bungu; 5°03'S, 38°24'E; Jul. 1912 • M.A. Mwangoka 26 (MO); Korogwe District, (T3) West Usambara Mountains, riverine forest patch in Kieti Village; 5°03'21'S, 38°24'17'E; alt. 1025 m; 31 May. 2002 • M.A. Mwangoka 6168 (MO); Muheza District, East Usambaras mountains, Kiwanda village Forest (Mpangamanyoka forest). W part of the forest; 5°02'32'S, 38°43'38'E; alt. 310 m; 05 Apr. 2009 • T.L.P. Couvreur 21 (DSM, K, MO, WAG); Muheza District, on road 200 m before arriving to Kisiwani village; 5°06'28.8'S, 38°40'55.2"E; alt. 300 m; 12 Nov. 2006.
Uvariodendron schmidtii
Type
Kenya – Coast • W.R.Q. Luke 3087 (holotype: EA! (EA000008814), isotypes: K!, MO, US); Kwale District, Shimba hills, Longo-Magandi; 4°14'S, 39°25'E; alt. 380 m; 20 Apr. 1992.
Description
Tree 10–12 m tall, D.B.H. unknown; young branches sparsely pubescent to glabrate, old branches glabrous; leaf bud ‘eragrostiform’, composed of 5–7, ca. 10 mm long, 10 mm wide, distichous, longitudinally folded, velutinous scales. Petiole 4.5–7 mm long, 1.5–2 mm wide, sparsely puberulent to glabrate. Leaf lamina 159–188 mm long, 49–71 mm wide, length:width ratio 2.4–3.3, narrowly elliptic to elliptic, coriaceous, base acute to decurrent, apex attenuate to acuminate, acumen 16–20 mm long; surface above glabrous, surface below sparsely pubescent to glabrate when young, glabrous when old; midrib impressed above, raised below, glabrous above, pubescent to glabrous when young, glabrous when old below; secondary veins 10–14 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and branches, composed of 1–2 flowers. Flower pedicel 10–15 mm long, 2.5 mm in diameter, densely velutinous. Flowers bisexual, buds globose, subsessile to pedicellate, ca. 6 mm high, ca. 7 mm in diameter, pubescent. Bracts 1 to 2 between the lower 20–70% of the length of the pedicel, upper bract ca. 5 mm long, ca. 10 mm wide, depressed ovate, adpressed, semi-clasping the pedicel, velutinous outside, glabrous inside. Sepals 3, 5.5–7 mm long, 7–9 mm wide, fused at base for more than 50% of the length, forming a ring around the fruit pedicel, densely velutinous to velutinous outside, glabrous inside, brown outside, green inside. Outer petals 3, 11–12 mm long, 9–11 mm wide, length:width ratio 1.1–1.3, broadly obovate, velutinous to densely velutinous outside, glabrous inside, brown outside, cream with a bright reddish-pink streak at base inside. Inner petals 3, ca. 10 mm long, 8–9 mm wide, length:width ratio 1.1–1.3, broadly obovate, connivent at apex over ca. 50 % of the petal length, densely velutinous outside, glabrous inside, orangish-brown with cream margins and a bright reddish-pink streak at base outside, cream with a bright reddish-pink streak at base inside. Stamens 500 to 700, length and shape unknown. Carpels ca. 7, ca. 1.5 mm long, ca. 1 mm wide, velutinous, free; stigma coiled. Fruiting pedicel ca. 16 mm long, ca. 2.5 mm in diameter, pubescent. Monocarps 3 to 5, ca. 32 mm long, ca. 20 mm wide, length:width ratio ca. 1.6, rounded to ellipsoid, longitudinally ridged, sparsely pubescent, green turning orange; sessile, stipe ca. 0 mm long. Seeds unknown.

Uvariodendron schmidtii Q.Luke, Dagallier & Couvreur A branch with leaves, upper side B detail of branch apex, note the ‘eragrostiform’ buds C two-flowered inflorescence, side view, bottom flower has no petals, note the sepals fused in A cup-shaped calyx D flower, top view E loose monocarps, side view. A, B, E Luke 4717 C, D unknown material. Photos A, B, E Léo-Paul Dagallier C, D Quentin Luke.
Distribution
Endemic to Somalia-Masai Region. Only known from one locality in Kenya: the Shimba Hills.
Habitat and ecology
Lowland rain forest. Soil: deep red. Altitude: 380–450 m a.s.l.
Phenology
Flowers collected in September and October. Fruits collected in April.
Notes
This species differs from other Uvariodendron species by its flowers that are small (petals < 13 mm long), velutinous, on a 10–15 mm long pedicel, with fused sepals forming a ring around the fruit pedicel, and fewer than 10 carpels.

Preliminary conservation status
Following IUCN criterion D, this species has been assigned a preliminary status of Vulnerable VU (
Additional specimens examined
Kenya – Coast • S.A. Robertson 7556 (EA, K, WAG); Kwale District, Shimba Hills NR, Longomandi forest; 4°14'S, 39°25'E; alt. 450 m; 04 Jun. 2005 • W.R.Q. Luke 2919 (EA); Kwale District, Shimba hills, Longo-Magandi; 4°14'S, 39°25'E; alt. 390 m; 15 Oct. 1991 • W.R.Q. Luke 4717 (EA, P); Kwale District, Shimba hills, Longo-Magandi; 4°14'S, 39°25'E; alt. 380 m; 12 Sep. 1997.
Uvariodendron usambarense
Type
Tanzania – Tanga • G. Scheffler 226 (lectotype, here designated: EA! (EA000002471); isolectotypes: A! (A00039714), BM! (BM000554074), E! (E00704857, E00704858), EA! (EA000002470), P! (P00363394); B sheet lost or destroyed), E Usambara Mts., Derema; 5°05'S, 38°38'E; 26 Mar. 1900.
Description
Tree 8–30 m tall, D.B.H. 17–30 cm; young branches pubescent to glabrous, old branches glabrous; the bark is dark on young individuals, peels on old individuals letting appear a pinkish-grey layer. Petiole 5–15 mm long, 1.5–6 mm wide, pubescent to glabrous. Leaf lamina 283–564 mm long, 79–180 mm wide, length:width ratio 2.4–3.9, obovate to oblong to elliptic, coriaceous, base rounded to subcordate, apex acuminate, acumen 5–18 mm long; surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 18–30 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Inflorescences borne on trunk and old branches, composed of 1 flower. Flower pedicel unknown. Flowers bisexual, buds ovoid, sessile, ca. 10 mm high, ca. 6 mm in diameter, pubescent. Only old rotten flower seen. Bracts unknown, 1 to 3 bracts scars observed on fruiting pedicel. Sepals 3, length, shape and indumentum unknown. Outer petals 3, ca. 20–30 (?) mm long, ca. 10–20 (?) mm wide (measures taken on old and rotten flower), length:width ratio ovate, greenish yellow to brown with dark red streak from base up to 50% of the petal length inside. Inner petals 3, length, shape, indumentum and color unknown. Stamens unknown. Carpels unknown. Fruiting pedicel 12–22 mm long, 4–8 mm in diameter, pubescent to glabrous. Monocarps 1 to 17, 50–89 mm long, 18–27 mm wide, length:width ratio 1.8–3.6, cylindrical, pubescent to sparsely pubescent, pale yellowish green (unripe ?); stipe 1–5 mm long, 6–10 mm wide, pubescent to glabrate. Seeds 16–25 per monocarp, biseriate, 20 to 25 mm long, 12–14 mm wide.

Distribution
Endemic to Somalia-Masai Region. Tanzania: Usambaras mountains and Udzungwa mountains.
Habitat and ecology
Submontane to montane rain forest. Altitude: 961–1000 m a.s.l.
Phenology
Flowers collected in August and November. Fruits collected from March to November.
Vernacular names
Tanzania: ‘Mkenene’ in Kinguru (Greenway 8628, Swynnerton 1110), ‘Mkomboa’ in Kisamba (Mwangoka 1593), ‘Ntambileni’ in Kinguru (Mwangoka 3272).
Notes
This species resembles Ud. calophyllum, Ud. connivens, Ud. fuscum and Ud. molundense in having great obovate to elliptic to oblong leaves (28–57 cm), but can be differentiated in having rounded to subcordate leaf bases (vs. acute to rounded), and a dark bark when young, peeling off when old, letting appear a pinkish-grey layer.
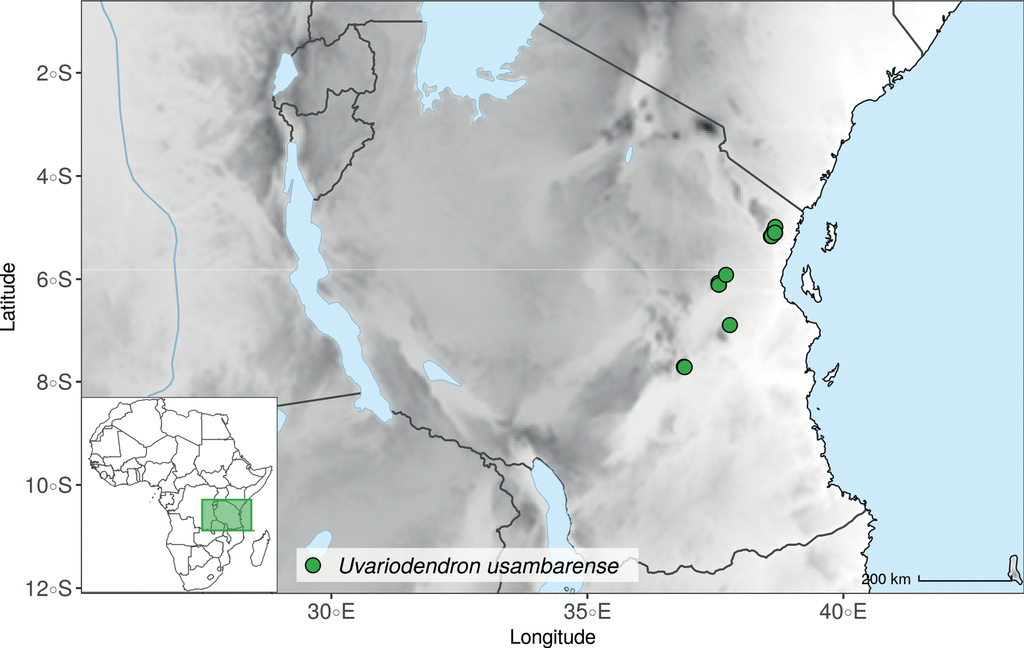
Preliminary conservation status
A previous assessment, which needs updating, listed this species as Endangered EN under criteria B1ab(iii) (Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project 2009d). Here the EOO is estimated at 13,513 km2 and AOO at 44 km2. It is known from 14 occurrences in Tanzania, situated in East Usambara Mountains, Uluguru Mountains and Udzungwa Mountains. Following IUCN criterion B, it would be assigned a preliminary conservation status of Endangered EN based solely on AOO, but no other subcriteria is met (b or c). Most of these occurrences are situated in protected areas such as the Udzungwa Mountains National Park and the Amani Nature Reserve. The future of this species relies on the future of these protected areas. It is thus assigned a preliminary updated conservation status of Near Threatened NT.
Additional specimens examined
Tanzania – Morogoro • A.R. Marshall 1100 (K), Msolwa – PSP9. Udzungwa National Park. udzungwa Mountains; 7°42'30.36'S, 36°53'06.73'E; alt. 1124 m; 05 Aug. 2007 • B.J. Harris 4610 (DSM, K); Morogoro Rural District, Uluguru Mtns. near Kinole; 6°54'S, 37°47'E; alt. 900 m; 30 Apr. 1970 • C.F.M. Swynnerton 1110 (K); Morogoro Rural District, S Nguru Forest Reserve, Manyangu Forest, Liwale River; 6°07'S, 37°34'E; Feb. 1920 • M.A. Mwangoka 3272 (MA, MO); Morogoro Rural District, Nguru Mountains, Nguru South Forest Reserve, along valley between Mafuta and Ubili villages; 6°04'34'S, 37°33'33'E; alt. 555 m; 12 Aug. 2004 • P.J. Greenway 8628 (BM, K); Morogoro Rural District, S Nguru Forest Reserve, Manyangu Forest, Liwale River; 6°07'S, 37°34'E; alt. 760 m; 21 Aug. 1951 • W.R.Q. Luke 8737 (EA, K, NHT), Pt 303, Camp 244; 7°43'S, 36°54'E; alt. 900 m; 06 Jun. 2002 • Y.S. Abeid 2666 (EA, MO); Mvomero District, Kanga Forest Reserve, Submontane Forest, Work Unit 3, Base Camp; 5°55'S, 37°42'28'E; alt. 1140 m; 23 Mar. 2006 – Tanga • A.L. Borhidi 82031 (DSM, K); Lushoto District, Amani-Sigi Forest Reserve at the Sigi headwaters S of Amani near Kwamkoro sawmills; 5°10'S, 38°36'E; alt. 900 m; 19 Feb. 1982 • F.M. Mbago 3707 (DSM); Lushoto District, Amani, above Kisiwani village; 5°06'53.38'S, 38°39'16.51'E; alt. 961 m; 16 Oct. 2015 • I. Rajabu Hizza 185 (MO); Muheza District, T3. Amani Nature Reserve, phenology transect P-T 2, Kisiwani; 5°06'S, 38°40'E; alt. 400 m; 17 Nov. 1998 • J.C. Lovett 816 (DSM, K, MO); Muheza District, East Usambara mountains, Kwamkoro Forest Reserve. Area currently being logged by Sikh sawmills; 5°10'S, 38°35'E; alt. 950 m; 11 Jun. 1986 • L.-P.M.J. Dagallier 34 (BR, DSM, K, MO, MPU, P, WAG); Muheza District, Amani Nature Reserve; 5°06'31.38'S, 38°39'45.44'E; alt. 687 m; 12 Nov. 2019 • L.B. Mwasumbi 112429 (DSM), Kwamkoro N. R; 5°10'S, 38°36'E; 08 Nov. 1998 • M.A. Mwangoka 1593 (MO); Muheza District, Kwezitu public forest along path to peak between Mkalamo and Gonja Subvillages; 4°59'17'S, 38°40'12'E; alt. 825 m; 04 Aug. 2000 • T.L.P. Couvreur 20 (DSM, MO, WAG); Muheza District, c. 1 km straight up from Kisiwani village (just before Amani Nature Reserve); 5°05'55.2'S, 38°39'40.2"E; alt. 1000 m; 11 Nov. 2006.
Uvariopsis
= Tetrastemma Diels, Bot. Jahrb. Syst. 38(3): 241 (1906).
= Thonnera De Wild., Ann. Mus. Congo Belge, Bot. sér. 5, 3(1): 86 (1909).
Type species
Uvariopsis zenkeri Engl.
Synoptic characters
Shrubs to trees with flowers invariably exhibiting two sepals, in combination with at least one of the two following characters (and generally with both of them): flowers unisexual (plant monoecious) and flowers exhibiting one whorl of four free or basally fused petals.
Description
Shrub to tree 1.5–30 m tall, D.B.H 1.5–39 cm; young branches pubescent to glabrous, old branches sparsely pubescent to glabrous. Petiole 1–9 mm long, 1–6 mm wide, pubescent to glabrous. Leaf lamina 73–615 mm long, 22–165 mm wide, length:width ratio 2.1–5, elliptic to oblong to obovate, papyraceous to coriaceous, base acute to decurrent to rounded to subcordate, apex acute to acuminate, acumen 0.5–30 mm long, surface above glabrous, surface below sparsely pubescent at base to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 5–26 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual or bisexual, male and female flowers similar or dimorphic, on same individuals (plant monoecious). Flower buds globose to long conical. Inflorescences borne in clumps of variable density on trunk, axillary or terminal, composed of 1 to 50 flowers. Peduncle inconspicuous to 2 mm long. Flower pedicel 0–430 mm long, 0.5–3 mm in diameter, pubescent to glabrous. Bracts 1 to 4, upper bract 0.5–4 mm long, 0.5–5 mm wide, triangular to broadly ovate, pubescent to sparsely pubescent outside, glabrous inside. Sepals 2, 0.7–18 mm long, 1–12 mm wide, triangular to broadly ovate, free to fused, pubescent to glabrous outside, glabrous inside, yellowish green to brown to dark red. Petals 3 to 4, 2.5–46 mm long, 1.5–22 mm wide, length:width ratio 0.9–5, elliptic to ovate, pubescent to glabrous outside, glabrous and soft to verrucose inside, cream to brown to dark red outside, cream to brown to dark red inside. Stamens 100 to 900, 0.1–2 mm long, 0.1–0.5 mm wide, anthers linear, connective prolongation truncate or absent. Carpels 3 to 280, 1–5 mm long, 0.5–3 mm wide, pubescent to glabrous, free; stigma 0.1–0.6 mm long, 0.2–1 mm wide, flat to coiled to globose, glabrous. Fruiting pedicel 2–400 mm long, 1–7 mm in diameter, pubescent to glabrous. Monocarps, 1–25, 10–85 mm long, 4.5–55 mm wide, length:width ratio 1.1–3.7, cylindrical, smooth to verrucose, straight to constricted between the seeds, pubescent to glabrous, dull olive green to red to brown, sessile to stipitate; stipe 0–14 mm long, 1–6 mm wide, pubescent to glabrous. Seeds 2–16 per monocarp, uniseriate to biseriate, 8–25 mm long, 5–15 mm wide, ellipsoid to semicircular.
Key to Uvariopsis species
| 1 | Largest leaves with lamina < 18 cm long, leaves with base acute to decurrent, flowers with petals < 16 mm long | 2 |
| – | Largest leaves with lamina > 18 cm long, leaves with base acute or rounded or subcordate or decurrent, flowers with petals ≥ or < 16 mm long | 6 |
| 2 | Leaves with base invariably decurrent, flowers bisexual with 3 to 6 carpels (Tanzania) | Up. bisexualis |
| – | Leaves with base acute to decurrent, flowers unisexual with more than 13 carpels | 3 |
| 3 | Young branches and petioles densely to sparsely pubescent, petals ovate to triangular, fused at base over ca. 50% of their length (Cameroon) | Up. zenkeri |
| – | Young branches glabrous or very sparsely pubescent, petals broadly ovate to ovate, free at base | 4 |
| 4 | Flowers with 13 to 20 carpels, fruits composed of 1 to 3 monocarps, not constricted between the seeds and pubescent to sparsely pubescent (West Africa) | Up. oligocarpa |
| – | Flowers with 20 to 40 carpels, fruits composed of 1 to 15 monocarps, strongly constricted between the seeds and glabrate to glabrous (Central and East Africa) | Up. congensis (5) |
| 5 | Shrub to tree 2–6 m tall, leaves elliptic to obovate with a length:width ratio between 2.1 and 3.1 (Central Africa) | Up. congensis var. congensis |
| – | Shrub to tree 7–15 m tall, leaves narrowly elliptic with a length:width ratio between 3 and 4 (Central and East Africa) | Up. congensis var. angustifolia |
| 6 | Larger leaves > 31 cm long, leaves with base rounded to subcordate or cordate | 7 |
| – | Larger leaves ≤ 31 cm long, leaves with base acute to decurrent, or acute to rounded to subcordate | 10 |
| 7 | Leaves emitting a strong lemon scent when crushed, flowering pedicel ≤ 2 mm long (Central Africa) | Up. citrata |
| – | Leaves not emitting lemon scent when crushed, flowering pedicel > 2 mm long | 8 |
| 8 | Female and male flowers with flowering pedicel ≤ 8 mm, petals narrowly ovate to linear, always free (Central Africa) | Up. bakeriana |
| – | Female flowers with pedicel ≥ 20 mm, petals ovate to narrowly ovate, free to fused | 9 |
| 9 | Largest leaves with lamina ≤ 38 cm long, female flowers with sepals ≥ 6 mm long, female and male flowers with petals pinkish to dark red inside and outside (Cameroon) | Up. submontana |
| – | Largest leaves with lamina > 38 cm long, female flowers with sepals ≤ 5 mm long, female and male flowers with petals cream to brownish outside and cream to pinkish inside (Central Africa) | Up. korupensis |
| 10 | Petals fused and female flowers with pedicel ≥ 8 cm | 11 |
| – | Petals free and female flower with pedicels < 20 cm, or petals fused and female flowers with pedicels ≤ 6.5 cm | 12 |
| 11 | Flowers with 3 petals, ≤ 40 carpels, monocarps stipitate (stipe 3–4 mm long) sparsely pubescent to glabrous with ca. 4 longitudinal ridges (Central Africa) | Up. congolana |
| – | Flowers with 4 petals, ≥ 50 carpels, monocarps sessile pubescent and verrucose (Central Africa) | Up. pedunculosa |
| 12 | Leaf base acute to decurrent | 13 |
| – | Leaf base acute and minutely cordate or acute to rounded to subcordate | 14 |
| 13 | Flowering pedicel ≤ 10 mm, ca. 20 carpels (Cameroon) | Up. etugeana |
| – | Flowering pedicel ≥ 14 mm, > 30 carpels (Tanzania) | Up. lovettiana |
| 14 | Leaf base acute and minutely cordate, inflorescence with peduncle ca. 2 mm long and 2 mm wide (Cameroon) | Up. dicaprio |
| – | Leaf base acute to rounded to subcordate, inflorescence with peduncle inconspicuous | 15 |
| 15 | Young branches and petiole tomentose to shortly tomentose (Central Africa) | Up. solheidii (16) |
| – | Young branches and petiole pubescent to glabrous | 17 |
| 16 | Male flowers with petals 7–10 mm long, with a l:w ratio of ca. 3; female flowers with pedicel 63–198 mm long and petals 9–17 mm long with a l:w ratio > 1.8; petals curved outward at anthesis (Central Africa) | Up. solheidii var. solheidii |
| – | Male flowers with petals 5–7 mm long, with a l:w ratio of ca. 2, female flowers with pedicel 20–110 mm long and petals 7.5–10 mm long with a l:w ratio > 1.8; petals straight at anthesis (Gabon) | Up. solheidii var. letestui |
| 17 | Flowers borne in clumps of 5 to 20 flowers on thickenings of the trunk, ≥ 100 carpels (Central Africa) | Up. dioica |
| – | Flowers borne in inflorescences of 1 to 2 flowers on the trunk or axillary or terminal, ≤ 50 carpels | 18 |
| 18 | Petals ovate with a l:w ratio ≥ 1.8, brownish red outside, cream inside; male flowers with pedicel ≤ 4 mm; female flower pedicel ≥ 70 mm (Angola) | Up. noldeae |
| – | Petals broadly ovate with a l:w ratio ≤ 1.4, pale greenish to cream outside, cream with purplish base inside; male flowers with pedicel ≥ 5 mm; female flower pedicel ≤ 65 mm (West Africa) | Up. guineensis (19) |
| 19 | Flowers with petals fused at base over 30–50% of their length and 20 to 50 carpels; male flowers with petals 8–17 mm long and 9–17 mm wide (West Africa) | Up. guineensis var. guineensis |
| – | Flowers with petals free and 10 to 20 carpels; male flowers with petals 7–10 mm long and 5–9 mm wide (Ghana) | Up. guineensis var. globiflora |
Species descriptions
Uvariopsis bakeriana
≡ Tetrastemma bakerianum Hutch. & Dalziel, Kew Bull. 1927: 153 (1927) & Fl. W. Trop. Afr. I. 57 (1927). Type. Nigeria – Cross River State • P.A. Talbot 1517 (holotype: K! (K000199043); isotype: BM! (BM000554076)); Calabar, Oban; 5°19'N, 8°34'E; 1912.
Description
Tree 3–7 m tall, D.B.H 1.8–8 cm; young branches pubescent to puberulent, old branches sparsely pubescent to glabrous. Petiole 2–6 mm long, 2–4 mm wide, pubescent to glabrous. Leaf lamina 150–340 mm long, 40–90 mm wide, length:width ratio (2.27) 3.3–5, oblong to obovate, coriaceous, base rounded to subcordate, apex acuminate, acumen 6–32 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 10–26 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers similar, on same individuals (plant monoecious). Flower buds long conical. Male and female inflorescences borne on trunk, composed of 1 to 2 flowers. Peduncle inconspicuous. Flower pedicel 2–8 mm long, 0.5–2 mm in diameter, pubescent. Bracts 1 (2) at base of the pedicel, 1–2 mm long, 1–4 mm wide, triangular, pubescent outside, glabrous inside. Sepals 2, 1–3.5 mm long, 1.5–3 mm wide, triangular, free, pubescent outside, glabrous inside, brown. Petals 4, 24–46 mm long, 3–8 mm wide, length:width ratio 4–10, narrowly ovate to linear, valvate, pubescent to sparsely pubescent outside, glabrous and highly verrucose inside, light red to dull yellowish brown outside, bright pink to dark pinkish red inside. Male flowers: stamens 400 to 600, 0.1–0.6 mm long, 0.1–0.3 mm wide, anthers linear, connective prolongation truncate or absent. Female flowers: carpels 15 to 40, 1.5–2.5 mm long, 1–1.5 mm wide, densely pubescent, free; stigma coiled. Fruiting pedicel 5–12 mm long, 2–2.5 mm in diameter, pubescent. Monocarps, 2–5, 21–50 mm long, 15–23 mm wide, length:width ratio 1.2–3.5, cylindrical, wrinkled, pubescent to glabrous, bright red, sessile to shortly stipitate; stipe up to 4 mm long, 1–3 mm wide, pubescent. Seeds 6–16 per monocarp, biseriate, 15–22 mm long, 8–12 mm wide, ellipsoid.

Uvariopsis bakeriana (Hutch. & Daltz.) Robyns & Ghesq A trunk with flowers B leaf, upper side C leaf base, lower side D leaf base, upper side E flowers borne on trunk F male flower, one petal removed, side view G male flower, top view H flower bud, side view I flower, side view J female flower, top view. A–D, F, H Couvreur 1000 E, I Couvreur 1045 G, J Couvreur 1015. Photos Thomas Couvreur.
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Cameroon, Equatorial Guinea (Bioko island), Nigeria.
Habitat and ecology
Lowland mature or old secondary rain forests. Soil: generally sandy. Altitude: 50–800 m asl.
Phenology
Flowers collected from January to April. Fruits collected from March to June.
Notes
This species resembles Up. citrata, Up. korupensis and Up. submontana in having large obovate leaves (up to 35 cm long), with rounded to subcordate bases. Up. bakeriana differs in having short flower pedicel (2–8 mm, vs. 0–2 mm in Up. citrata, 6–70 mm in Up. korupensis and 25–60 mm in Up. submontana), and with its petals being narrowly ovate to linear, with a length:width ratio comprised between 4 and 10 (vs. ca. less than 5 in Up. citrata and Up. submontana), free (vs. fused in Up. korupensis), and bright pinkish to deep red petals (vs. cream to brownish in Up. citrata and Up. korupensis) (Table

Uvariopsis bakeriana (Hutch. & Daltz.) Robyns & Ghesq A leaf, top view B detail of female flower, two petals removed C petal, inner view D carpel, side view, front view and detail of ovules E monocarp Uvariopsis dioica (Diels) Robyns & Ghesq F leaf, top view G male flowering pedicels and flowers H detail of male flower, two petals removed I stamen J female flowering pedicel K detail of female flower, all four petals removed L carpel, front view and detail of ovules M fruits. A–E from Brenan 9409 F–L from Keay 28066 M from Letouzey 4230. Drawings by Hélène Lamourdedieu, Publications Scientifiques du Muséum national d’Histoire naturelle, Paris.
The ‘large leaves group’: morphological comparison between Uvariopsis bakeriana, Uvariopsis citrata, Uvariopsis korupensis and Uvariopsis submontana. In bold: character useful to differentiate the species. l:w = length:width.
| Lamina | Flower pedicel | Sepals | Petals | Altitudinal range | |
|---|---|---|---|---|---|
| Uvariopsis bakeriana | 150–340 mm long | 2–8 mm long | Male and female flowers: 1–3.5 mm long, 1.5–3 mm wide, free | Male and female flowers: l:w ratio 4–10, narrowly ovate to linear, free, valvate, bright pink to dark pinkish red | 50–800 m above sea level |
| Uvariopsis citrata | 312–500 mm long, strong lemon scent when crushed | 0–2 mm long | Male and female flowers: 9–15 mm long, 4–6 mm wide, basely fused | Male and female flowers: l:w ratio ca. 3.5, ovate, greenish yellow | 60–300 m above sea level |
| Uvariopsis korupensis | 280–615 mm long | 6–70 mm long | Male flowers: 1–5 (7.5) mm long, 2–6.5 mm wide, basely fused | Male flowers: l:w ratio 2.5–7, narrowly ovate, fused at base | 90–160 m above sea level |
| Female flowers: 3–5 mm long, 4–5 mm wide, basely fused | Female flowers: petals more than 3 times longer than the sepals, l:w ratio 7–12, narrowly ovate, fused at base, cream to pinkish | ||||
| Uvariopsis submontana | 160–380 mm long | 24–60 mm long | Male flowers: 5–11 mm long, 6–12 mm wide, basely fused | Male flowers: l:w ratio 1.8–4.7, ovate to narrowly ovate, free to fused at base over 30% of their length | 900–1300 m above sea level |
| Female flowers: 6–8 mm long, 6–9 mm wide, basely fused | Female flowers: petals less than 3 times longer than the sepals, l:w ratio 5–9, narrowly ovate, fused at base, ovate to narrowly ovate, free to fused at base over 30% of their length, cream to pinkish |
Conservation status
This species has been assessed as Least Concern LC (

Additional specimens examined
Cameroon – Littoral • D.W. Thomas 4300 (K, MO), Korup National Park. Forest along footpath from Ndian River at PAMOL field 69 and transect P; 5°01'N, 8°50'E; alt. 50 m; 24 Jan. 1985 – South-West Region • C. Farron 7297 (P), South Bakundu Forest, Kendongi; 4°26'30.63'N, 9°20'43.88'E; 14 May. 1970 • C.F.A. Onochie 9305 (K), South Bakundu Forest; 4°24'06.26'N, 9°26'32.16'E; 12 Mar. 1948 • D.W. Thomas 1086 (K); Ndian, Korup Forest Reserve, Korup Reserve, path between transect P&Q; 4°59'N, 8°51'E; Mar. 1979 • D.W. Thomas 3210 (K, MO), forest in the Korup National Park; 5°03'N, 8°48'E; alt. 50 m; 28 Feb. 1984 • D.W. Thomas 3336 (MO), North-eastern corner of Korup National Park; near Baro village; 5°16'N, 9°11'E; alt. 200 m; 24 Mar. 1984 • D.W. Thomas 5606 (MO), along transect P, southern end of Korup National Park; 5°11'N, 8°51'E; alt. 50 m; 16 Feb. 1986 • D.W. Thomas 7835 (MO), Ajaman, near northern edge of Korup National Park; 5°20'N, 8°54'E; alt. 200 m; 22 May. 1988 • G.K. Gottsberger 100307/11 (ULM, WAG), c. 100 m from Banyang Mbo Research Station; 5°08'N, 9°30'E; 10 Mar. 2007 • J.P.M. Brenan 9305 (K), South Bakundu Forest; 4°29'N, 9°23'E; 12 Mar. 1948 • J.P.M. Brenan 9409 (BM, K), Banga, S. Bakundu F.R; 4°24'N, 9°27'E; 13 Mar. 1948 • M.R. Cheek 7234 (K, SCA, YA); Ndian, Mundemba, Korup National Park, nature trail near suspension bridge.; 4°59'N, 8°51'E; alt. 50 m; 01 Feb. 1995 • R.E. Gereau 5195 (MA, MO, WAG); Ndian, 2–2.5 km S of Six Cup Garri Creek; 5°02'N, 8°53'E; alt. 100 m; 06 Mar. 1993 • R.G. Letouzey 13841 (P), entre Etinkem et Nfaitok, 10 km N Nguti; 5°25'39.72'N, 9°25'14.26'E; 15 Jun. 1975 • S. Cable 2150 (K, YA); Ndian, Korup National Park, Ekundu Kundu, path to Ekon 1, 5 km from Ekundu-Kundu; 5°04'50.32'N, 8°54'29.81'E; alt. 300 m; 25 Apr. 1996 • S. Cable 2256 (K, YA); Ndian, Korup National Park, path from Ekundu Kundu to Faba, about 3 km; 5°02'53.41'N, 8°56'25.12'E; alt. 300 m; 27 Apr. 1996 • T.L.P. Couvreur 1000 (MPU, WAG, YA), Bayang Mbo Wildlife Sanctuary, after Mbu river; 5°21'09.69'N, 9°30'09.23'E; alt. 259 m; 25 Mar. 2016 • T.L.P. Couvreur 1015 (MPU, WAG, YA), Bayang Mbo Wildlife Sanctuary, after Mbu river; 5°21'22.04'N, 9°30'05.8'E; alt. 250 m; 26 Mar. 2016 • T.L.P. Couvreur 1045 (MPU, WAG, YA), Mount Cameroon National Park, on the Bomona trail, behind Bomona village, 10 km NW from Idenau; 4°17'41.96'N, 9°05'53.93'E; alt. 856 m; 03 Apr. 2016 • W. Mukete 6 (K, MO, SCA, YA), Kosse, along the path to the Reserve; 4°19'48.72'N, 9°02'52.08'E; 24 Apr. 1996. Equatorial Guinea – Bioko Sur • W.R.Q. Luke 11907 (K, MA), Moaba. Pt 140-141; 3°18'20.6'N, 8°39'33.8'E; alt. 8 m; 15 Mar. 2007. Nigeria – Cross River State • C.F.A. Onochie FHI36140X (K); Calabar, Oban Group Forest Reserve, Orem; 5°36'N, 8°35'E; 23 Jan. 1957 • J. Ntui 944 (MO), Oban Hills; 5°33'30'N, 8°34'12'E; alt. 600 m; 19 Dec. 1955 • M.C. Ejiofor FHI21870 (K); Calabar, Oban Group Forest Reserve; 5°36'N, 8°35'E; 24 Apr. 1952 • P.A. Talbot 1516 (BM); Calabar, Oban; 5°19'N, 8°34'E; 1912 • P.A. Talbot 446 (BM), Oban; 5°13'23.28'N, 8°33'06.9'E; 1909 • P.W. Richards 5166 (K, K, P); Calabar, Oban; 5°13'27.54'N, 8°33'10.42'E; alt. 244 m; 13 Mar. 1955 – Ogun State • A.P.D. Jones FHI17275 (P), c. 1/2 ml W. of Oaho Enclave, E.B. 34 Line 7. Road trace of U.A.C; 7°16'N, 4°12'E; 05 Apr. 1946.
Uvariopsis bisexualis
Type
Tanzania – Iringa • J.C. Lovett 233 (holotype: K! (K000199040)); Iringa Rural District, Udzungwa Mts., Sanje; 7°46'S, 36°54'E; alt. 1480 m; 02 Dec. 1983.
Description
Tree ca. 10 m tall, D.B.H unknown; young branches pubescent, old branches glabrous. Petiole 3.5–5 mm long, ca. 2 mm wide, sparsely pubescent to glabrous. Leaf lamina 135–152 mm long, 37–48 mm wide, length:width ratio ca. 3, elliptic to oblong, coriaceous, base decurrent, apex attenuate, surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins ca. 12 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers bisexual. Flower buds ovoid to conical. Inflorescences borne on branches, composed of 1 flower. Peduncle inconspicuous. Flower pedicel 12–20 mm long, ca. 1 mm in diameter, pubescent. Bracts 1 at base to towards the lower half of the pedicel, ca. 1 mm long, ca. 1 mm wide. Sepals 2, 1.6–2.5 mm long, 2.2–2.5 mm wide, broadly ovate, free, pubescent outside, glabrous inside, color unknown. Petals 4, 12–14 mm long, 4–8 mm wide, length:width ratio 3–3.5, narrowly ovate, sparsely pubescent outside, glabrous inside, cream outside, cream with red base inside. Stamens numerous (exact number unknown), 0.5–0.7 mm long, ca. 0.5 mm wide, anthers linear, connective prolongation truncate or absent. Carpels 3 to 6, 2–4 mm long, 1–1.4 mm wide, glabrate at base to glabrous, free, ovules 22–24 per ovary, biseriate. Fruits unknown.

Uvariopsis bisexualis Verdc A flowering branch B flower bud, side view C sepal, outer view D flower, two petals removed, side view E detail of receptacle, some stamens fallen, side view F stamen, upper view G stamen, lower view H carpel, outer view J carpel, inner view K stigma L longitudinal section of carpel M transversal section of ovary. A–M from Lovett 233 (type). Drawings by Maureen Church, from
Distribution
Endemic to Somalia-Masai Region. Known from only one locality in Tanzania: Sanje in the Udzungwa mountains.
Habitat and ecology
Montane rain forests. Altitude ca. 1480 m asl.
Phenology
Flowers collected in December.
Notes
This species resembles Up. congensis, Up. oligocarpa and Up. zenkeri in having elliptic leaves generally less than 16 cm long with decurrent base. It differs from all the other Uvariopsis in having bisexual flowers and less than 6 carpels (vs. more than 10 carpels) (Table

Morphological comparison between Uvariopsis bisexualis, Uvariopsis congensis, Uvariopsis oligocarpa and Uvariopsis zenkeri. In bold: character useful to differentiate the species.
| Young branches indumentum | Flowers | Carpels number | Fruits | |
|---|---|---|---|---|
| Uvariopsis bisexualis | pubescent | bisexual | 3 to 6 | unknown |
| Uvariopsis congensis | sparsely pubescent to glabrous | unisexual | 20 to 40 | 1 to 15 monocarps, cylindrical, smooth to lumpy, strongly constricted between the seeds, glabrate to glabrous, green to orange to red when ripe (black when dry), sessile to stipitate with stipe 0–7 mm long |
| Uvariopsis oligocarpa | glabrous | unisexual | 13 to 20 | 1–3 monocarps, cylindrical, smooth, pubescent to sparsely pubescent, green to yellow to orange when ripe (orange-brown when dry), sessile to very shortly stipitate with stipe 0–1 mm long |
| Uvariopsis zenkeri | densely pubescent to pubescent | unisexual | 13 to 22 | 1–3 monocarps, cylindrical, slightly veined and constricted between the seed when dried, tomentose, brown, subsessile with stipe ca. 1 mm long |
Conservation status
This species has been assessed as Endangered EN under criteria B1ab(iii) (Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project 2009e). It is known from a single specimen collected more than 30 years ago in the Udzungwa Mountains National Park in Tanzania.
Uvariopsis citrata
Type
Gabon – Estuaire • T.L.P. Couvreur 1143 (holotype: WAG! (no barcode); isotypes: LBV, P!), Monts de Cristal National Park, Mbé sector, 800 m from Kinguélé ANPN camp, near bridge; 0°27'50.18'N, 10°16'42.82'E; 14 Jun. 2016.
Description
Tree 4–10 m tall, D.B.H 3–10 cm; young branches pubescent, old branches glabrous. Leaves strong lemon scent when crushed. Petiole 4–8 mm long, 3–5 mm wide, pubescent to glabrous. Leaf lamina 312–500 mm long, 88–120 mm wide, length:width ratio 3.5–4.3, elliptic to obovate, coriaceous, base subcordate, apex acuminate, acumen 20–30 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 17–19 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers similar, on same individuals (plant monoecious). Flower buds ovoid to conical. Male and female inflorescences borne on trunk, sparsely spaced mostly towards the lower half of the trunk, composed of 1 to 2 sessile flowers. Peduncle inconspicuous. Flower pedicel 0–2 mm long, 1–2 mm in diameter, densely pubescent. Bracts up to 3 at base, upper bract 1–2 mm long, ca. 4 mm wide, broadly ovate, pubescent outside, glabrous inside. Sepals 2, 9–15 mm long, 4–6 mm wide, narrowly ovate, basally fused, enclosing the petals in bud, densely pubescent outside, densely pubescent to glabrous toward base inside, brown. Petals 4, 7–15 mm long, 5–8 mm wide, length:width ratio ca. 3.5, ovate, pubescent outside, glabrous inside, brownish to greenish yellow outside, greenish yellow inside. Male flowers: stamens number unknown, ca. 0.5 mm long, anthers linear, connective prolongation truncate. Female flowers: carpels 60, 4–5 mm long, ca. 0.5 mm wide, densely pubescent, free; stigma coiled. Fruits unknown.

Uvariopsis citrata Couvreur & Niang A trunk with flower buds B leaves, upper side C leaf base, lower side D lear apex, upper side E leaf base, upper side F female flower bud, two petals removed, side view G male flower bud, one petal removed, side view H flower buds on trunk. A–E, H Couvreur 1143 (type) F, G Couvreur 1126. Photos Thomas Couvreur.
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Cameroon, Gabon.
Habitat and ecology
Lowland mature or old secondary rain forests, near streams or in periodically inundated forest. Altitude: 60–300 m asl.
Phenology
Flowers collected from March to June.
Uses
Young leaves are used to wrap fish while cooking to give it an aromatic taste (Letouzey 9017).
Notes
This species resembles Up. bakeriana, Up. korupensis and Up. submontana in having large obovate leaves (more than 31 cm long), with rounded to subcordate bases. Up. citrata is unique within the genus by its leaves emitting a strong lemon scent when crushed (Table

Conservation status
This species is known from one locality in Gabon situated in the Monts de Cristal National Park, and two localities in Cameroon both situated in the Campo-Ma’an National Park. It has been previously assessed as Data Deficient based on the Gabonese population (
Additional specimens examined
Cameroon – South Region • G.P. Tchouto Mbatchou 2869 (KRIBI, WAG, YA), Campo-Ma'an area, Bibabimvoto; 2°12'48'N, 10°00'51'E; alt. 60 m; 13 May. 2000 • R.G. Letouzey 9017 (P), 15 km au SSE de Zingui (soit à 50 km au SE de Kribi), feuille IGN 1/200000 Nyabessan; 2°37'52.24'N, 10°15'58.54'E; 14 Mar. 1968. Gabon – Estuaire • T.L.P. Couvreur 1126 (LBV, WAG), Monts de Cristal National Park, 800 m from Kinguélé ANPN camp, near bridge; 0°27'50.18'N, 10°16'42.82'E; 11 Jun. 2016.
Uvariopsis congensis
Type
Democratic Republic of the Congo – Kasai-Oriental • E. van der Kerkhoven s.n (lectotype, here designated: BR! (BR0000008824318)); Lusambo, Saint-Trudon [=Mombelaye]; 5°04'00.23'S, 23°28'59.88'E; alt. 475 m; 16 Aug. 1913.
Description
Shrub to tree 2–15 m tall, D.B.H 3–39 cm; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 2–5 mm long, 1–2.5 mm wide, glabrous. Leaf lamina 73–177 mm long, 22–77 mm wide, length:width ratio 2.1–4, elliptic to narrowly elliptic to obovate, papyraceous to coriaceous, base acute to decurrent, apex attenuate to acuminate, acumen 5–18 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 7–12 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers similar, on same individuals (plant monoecious). Flower buds globose. Male and female inflorescences borne on old branches or axillary, composed of 1 to 2 flowers. Peduncle inconspicuous. Flower pedicel 3–11 mm long, 0.5–1 mm in diameter, pubescent to glabrous. Bracts 1 to 4 at base and one towards the lower half of pedicel, upper bract ca. 1 mm long, ca. 1 mm wide, triangular, pubescent outside, glabrous inside. Sepals 2, 0.7–1.5 mm long, 1.5–2 mm wide, broadly ovate, free, pubescent outside, glabrous inside, brown. Petals 4, 2.5–8 mm long, 1.5–5 mm wide, length:width ratio 1–1.7, ovate, free, pubescent to glabrate outside, glabrous inside, cream to pale orange outside, cream to yellow inside. Male flowers: stamens 300 to 400, ca. 0.5 mm long, ca. 0.2 mm wide, anthers linear, connective prolongation truncate or absent. Female flowers: carpels 20 to 40, 1.5–2 mm long, 0.5–1 mm wide, pubescent, free; stigma 0.2–0.5 mm long, ca. 0.5 mm wide, globose to flat. Fruiting pedicel 5–15 mm long, 1–3 mm in diameter, pubescent to glabrous. Monocarps, 1–15, 13–45 mm long, 7–19 mm wide, length:width ratio 1.3–3.1, cylindrical, smooth to lumpy, strongly constricted between the seeds, apex rounded to slightly acuminate, glabrate to glabrous, green to orange to red when ripe (black when dry), sessile to stipitate; stipe 0–7 mm long, 1–3 mm wide, glabrous. Seeds 2–11 per monocarp, uniseriate to biseriate, 9.5–14 mm long, 7–10 mm wide, ellipsoid.

Uvariopsis congensis Robyns & Ghesq A leaf, lower side B female flower, all carpels fallen, top view C male flower, stamens on the top of the receptacle fallen, semi-top view D male flower, dry, side view E female flower, three petals removed, side view F fruit, fresh, top view G fruit, dry, side view. A, F Texier 2307 B, C, G Lachenaud 1384 D Wieringa 660 E Tisserant 1363. Photos A, F Nicolas Texier (CC BY-NC-ND 3.0) B, C Olivier Lachenaud (CC BY-NC-ND 3.0) D, E, G Léo-Paul Dagallier.
Distribution
Element of Lower Guinean Domain and Congolia Domain of the Guineo-Congolian Region and Zambezian Region: Angola, Cameroon, Central African Republic, Democratic Republic of the Congo, Gabon, Kenya, Republic of the Congo, Uganda.

Habitat and ecology
Lowland and premontane mature or secondary rain forests, often in periodically inundated forest. Altitude: 18–1000 m asl.
Phenology
Flowers and fruits collected all year.
Vernacular names
Central African Republic: ‘Ikuta’ (Tisserant 1504) and ‘Molo-Nzange’ (Tisserant 2136) in Lissongo.
Notes
This species resembles Up. oligocarpa and Up. zenkeri in having elliptic leaves less than 18 cm long at maximum, with a decurrent base. It differs from Up. oligocarpa and Up. zenkeri in having more carpels (20 to 40 vs. 13 to 20 in Up. oligocarpa and 13 to 22 in Up. zenkeri) and glabrous monocarps strongly constricted between the seeds (vs. pubescent and not constricted in Up. oligocarpa and tomentose and slightly constricted in Up. zenkeri) (Table
Conservation status
This species is widespread, distributed from Cameroon to Kenya. It has been previously assessed as Least Concern LC (
Uvariopsis congensis var. angustifolia , var. nov.
Type
South Sudan – Unknown major area • I. Friis 735 (holotype: K! (no barcode)), Talanga; 4°01'N, 32°45'E; alt. 1300 m; 07 Dec. 1980.
Diagnosis
Up. congensis var. angustifolia differs from the type variety in being trees 7–15 m high (vs. shrubs to trees 2–6 m high) and having narrowly elliptic leaves with a length:width ratio between 3 and 4 (vs. elliptic to obovate leaves with a length:width ratio between 2.1 and 3.1).
Description
Tree 7–15 m tall. Leaf lamina 73–166 mm long, 22–46 mm wide, length:width ratio 3–4, narrowly elliptic.
Distribution
Element of the Congolia Domain of the Guineo-Congolian Region and the Somalia-Masai Region: Democratic Republic of the Congo, Kenya, Uganda.
Habitat and ecology
Lowland and premontane mature or secondary rain forests. Altitude: 470–1000 m asl.
Notes
The variety Uvariopsis congensis var. angustifolia was created to acknowledge the clear morphological variation exhibited by several specimens (mainly from East Africa), being taller trees and having narrower leaves than the other specimens (mainly from Central Africa), although this morphotype is not monophyletic (see Fig.
Additional specimens examined
Democratic Republic of the Congo – Orientale • J.-P.A. Lebrun 2463 (P), Likati Uele; 3°32'25'N, 24°02'11'E; Mar. 1931 • J.-P.A. Lebrun 2619 (P); Buta, Buta-Uele; 2°48'16.32'N, 24°44'59.68'E; Mar. 1931 • J.L.P. Louis 9562 (K); Isangi, Yalulia, à 20 km à l'Est de Yangambi; 0°40'N, 24°38'E; alt. 470 m; 30 May. 1938. Kenya – Nyanza • B. Verdcourt 1695 (K); North Kavirondo, Kakamega Forest Station, By roadside leading to Yala River from Junction of main road with road leading to Forest House section. Kakamega Forest; 0°14'N, 34°51'E; 10 Dec. 1956 • R.B. Faden 70/34 (EA, K); North Kavirondo, Kakamega Forest, Kibiri Block, S. side of Yala River; 0°11'N, 34°52'E; alt. 1550 m; 21 Jan. 1970. Uganda – Buganda • B.T. Styles 236 (K); Mengo, Mabira C.F.R., coupe 71; 0°29'03.48'N, 32°59'05.64'E; alt. 1100 m; 23 Nov. 1962 – Central • R.A. Dümmer 5591 (K), Mulange; 0°30'28.76'N, 33°02'39.79'E; Oct. 1922 – Western Province • B.T. Styles 122 (K); Masindi District, Budongo Forest Reserve, Waibira Block Compt. 19; 1°43'N, 31°32'E; alt. 1097 m; 04 Oct. 1962 • C.M. Harris 167 (K); Masindi District, Budongo Forest Reserve; 1°43'N, 31°32'E; Nov. 1932 • D.L.N. Hafashimana 13 (K); Masindi District, Budongo Forest Reserve, Kaniyo – Pabidi beat; 1°43'N, 31°32'E; alt. 1010 m; Feb. 1996 • H.A. Lindeman 534 (K); Masindi District, Budongo Forest Reserve; 1°43'N, 31°32'E; Mar. 1939 • J.G. Myers 13647 (K); Masindi District, Budongo Forest Reserve; 1°43'N, 31°32'E; 20 Dec. 1940 • M.T. Dawe 484 (K); Toro, Toro at Isungu; 0°28'59.88'N, 30°19'00.12'E; alt. 1500 m; 06 Sep. 1905 • S. Paulo 581 (K); Kabarole, Fort Portal Forest Reserve; 0°40'N, 30°16'E; 18 Jul. 1960 • W.J. Eggeling 2291 (EA, K); Masindi District, Budongo Forest Reserve; 1°43'N, 31°32'E; Nov. 1935 • W.J. Eggeling 3154 (K); Toro, Kibale Forest; 0°30'N, 30°24'E; Aug. 1936 • W.J. Eggeling 3357 (K); Masindi District, Budongo Forest Reserve; 1°43'N, 31°32'E; Jun. 1937. Zambia – North-Western • F. White 3334 (BM, K, MO); Mwinilunga, west of Kalene Hill Mission; 11°10'02.24'S, 24°09'54.84'E; 22 Sep. 1952.
Uvariopsis congensis var. congensis
Description
Shrub to tree 2–6 m tall. Leaf lamina 88–177 mm long, 30–77 mm wide, length:width ratio 2.1–3.1 (3.6), elliptic to obovate.
Distribution
Element of the Lower Guinean Domain and Congolia Domain of the Guineo-Congolian Region and the Zambezian Region: Angola, Cameroon, Central African Republic, Democratic Republic of the Congo, Gabon, Republic of the Congo.
Habitat and ecology
Lowland and premontane mature or old secondary rain forests, often in periodically inundated forest. Altitude: 18–1000 m asl.
Additional specimens examined
Angola – Lunda Norte • I.A. Darbyshire 759 (K), Lovue River near Capaia village; 8°20'17'S, 20°14'25'E; alt. 960 m; 30 Mar. 2013 • J. Gossweiler 13802a (K), Sector fitocologico de Nordeste de Lunda, circunscrição de Chitato. Dundo, Luachimo; 7°24'S, 20°50'E; alt. 700 m; 05 Nov. 1946 • J. Gossweiler 14071 (B, P); 7°54'S, 21°21'E; alt. 700 m; 12 Jun. 1948. Cameroon – Central Region • W.J.J.O. de Wilde 1914 (P, WAG), C. 30 km South of M'Balmayo; 3°20'N, 11°30'E; 13 Feb. 1964 – East Region • D.J. Harris 1512 (K, MO, P), West bank of Sangha River; 2°21'N, 16°09'E; alt. 350 m; 01 Nov. 1988 • F.J. Breteler 2812 (K, P, WAG), Near Bimba, bank of Doumé river, 40 km SW. of Batouri; 4°10'N, 14°07'E; alt. 580 m; 15 Apr. 1962 • J.F. Villiers 683 (P), 2 km N du confluent Ngoko-Malapa, 5 km E de Moloundou; 2°03'17.9'N, 15°11'01.31'E; 21 Apr. 1971 • R.G. Letouzey 10641 (P), Bordure de la Sangha au Sud de Lidjombo (près ile Libongo) à 110 km au N. de Ouesso (feuilles IGN 1/200.000 Moloundou); 2°40'N, 16°05'E; 10 Apr. 1971 • R.G. Letouzey 12086 (K, P, WAG), près Ndongo, à 45 km WNW de Moloundou (Feuille IGN 1/200.000 SOUANKE); 2°05'N, 14°52'E; 15 Mar. 1973 • R.G. Letouzey 4755 (K, P), Rives de la Doumé près Bimba 40 km SW de Batouri – feuille IGN 1/200.000 Batouri-Berberati; 4°10'32.91'N, 14°06'32.96'E; 15 Apr. 1962 • R.G. Letouzey 5491 (K, P), a 11 km au SSW de Koso (village situé à 60 km au SSW de Batouri); 3°50'23.13'N, 14°07'33.62'E; 25 Jul. 1963 – South Region • R.G. Letouzey 10024 (P), Rives du Ntem près du confluent de la Kye, 16 km ESE d'Ambam; 2°18'02.17'N, 11°23'15.84'E; 07 Feb. 1970 • R.G. Letouzey 9915 (P), Bord rivière Nlobo, près Ngomebae, 70 km ESE d'Ebolowa sur route Mvangan; 2°43'N, 11°45'E; 24 Jan. 1970 – South-West Region • A. Staudt 556 (K), Johann – Albrechtshöhe, Johann – Albrechtshohe, Kumba area; 4°10'N, 9°12'E; 1896. Central African Republic – Basse-Kotto • C. Tisserant 1363 (P), Galerie Riv. Moku Sikumdu 10 km N.O. Alindas; 5°10'24.36'N, 21°05'07.74'E; 19 Nov. 1927 – Lobaye • Équipe C. Tisserant 1504 (BM, BR, K, P); Mbaïki, Boukoko; 3°54'N, 17°56'E; 15 Jun. 1949 • Équipe C. Tisserant 2136 (BR, P); Mbaïki, Boukoko; 3°54'N, 17°56'E; 16 Jun. 1951 • Équipe C. Tisserant 2198 (BM, BR, P); Mbaïki, Boukoko; 3°54'N, 17°56'E; 17 Aug. 1951 • Équipe C. Tisserant 2241 (BM, BR, K, P, P); Mbaïki, Boukoko; 3°54'N, 17°56'E; 13 Sep. 1951 • F.J. Badré 239 (B, P); Mbaïki, Mbaiki, Bord de la Lobaye – Forêt 30 km S de Mbaïki; 3°38'25.5'N, 18°00'57.02'E; 06 Nov. 1968. Democratic Republic of the Congo – Bas-Congo • F.H.E.A.W. Robyns 233 (BR); Madimba, Kisantu, Station Rwé droite, Inkisi; 5°08'S, 15°05'E; 10 Jul. 1925 • J. Gillet 22 (BR (BR0000014588372)); Madimba; 1899 – Maniema • C.E.N. Ewango 2664 (MO), Territoire de Kailo, PArc National de la Lomami. Localité Katopa (Camp); 2°46'52'S, 25°06'34'E; alt. 458 m; 21 Mar. 2015 – Orientale • J. Bokdam 3388 (KIS, WAG); Kisangani, 10 km W of Kisangani, near Lindi river; 0°33'N, 25°06'E; 19 Nov. 1971 • J.-P.A. Lebrun 2487 (P); Buta, Buta-Uele; 2°48'16.32'N, 24°44'59.68'E; Mar. 1931 • J.L.P. Louis 13423 (EA, K, MO, P); Isangi, Bassao (île), Un peu en aval de Liléko; 0°54'N, 24°12'E; alt. 470 m; 26 Jan. 1938 • M.D.J. Laurent 1618 (BR); Banalia, Bomaneh; 1°18'N, 23°47'E; 04 Mar. 1906 • R.G.A. Germain 4757 (K), à l'W de Basoko, embouchure de la Lombo; 1°14'14.83'N, 23°34'46.46'E; Feb. 1949 • R.G.A. Germain 4804 (P), Yankeleli, près d'Isangi; 0°44'31'N, 24°12'15'E; Apr. 1949 • R.G.A. Germain 50 (B, K, P); Isangi, village de Yafolo, rives gauche en amont d'Isangi; 0°46'46'N, 24°16'13'E; alt. 470 m; 21 Dec. 1939 • R.G.A. Germain 8207 (K, M); Isangi, île Esali III (en aval d'Isangi); 0°53'34.91'N, 24°13'08.13'E; Feb. 1953 • R.G.A. Germain 8769 (K); Isangi, Yabwesu-Ogeto (Bas Lomami); 0°38'N, 23°54'E; 05 Apr. 1956 – Unknown major area • M.D.J. Laurent s.n (BR); 1°19'N, 23°48'E; 04 Mar. 1906. Gabon – Moyen-Ogooué • T.O.B.E.B. Stévart 4523 (LBV, MO, P), Mabounié, along the Ngounié River; 0°49'59'N, 10°33'23'E; alt. 18 m; 11 May. 2012 – Ngounié • O.L.S. Lachenaud 1384 (BR, BRLU, LBV, MO, P, WAG), Mabounié, rive gauche de la Ngounié juste en aval du débarcadère; 0°48'40'N, 10°29'57'E; alt. 24 m; 15 Nov. 2013 • Ogooué-Lolo • F.J. Breteler 6647 (LBV, P, WAG), 4 km SW of Lastoursville, right side Ogooué R; 0°50'N, 12°41'E; 25 Sep. 1970 – Woleu-Ntem • G.M.P.C. Le Testu 9027 (BM, P), region de Bitam, Meyo kyè; 2°10'N, 11°22'E; Mar. 1933 • J.J. Wieringa 660 (WAG), c. 85 km N of Makokou, Minkébé district, Nsye valley; 1°22'N, 12°56'E; alt. 505 m; 27 Feb. 1990 • MINKébé Series W626 (K, LBV, MAKOK, MO, P, WAG), Minkébé area, river Nouna; 1°43'N, 12°51'E; 16 Dec. 1990. Republic of the Congo – Pool • A.J.B. Chevalier 4176 (P), Brazzaville; 4°16'S, 15°17'E; Jul. 1902.
Uvariopsis congolana
≡ Thonnera congolana
De Wild., Ann. Mus. Congo Belge, Bot. sér. 5, 3(1): 86 (1909). Type. Democratic Republic of the Congo – Equateur • F. Thonner 100 (holotype: BR! (BR0000008824394, BR0000008824202), sheet designated in
Description
Tree 1.5–10 m tall, D.B.H ca. 5 cm; young branches pubescent to glabrous, blackish when dry, old branches glabrous. Petiole 2–5 mm long, 1.5–4 mm wide, pubescent to glabrous. Leaf lamina 163–300 mm long, 44–110 mm wide, length:width ratio 2.4–4, (elliptic) oblong to obovate, papyraceous to coriaceous, base acute to rounded, apex attenuate to acuminate, acumen 2.5–22 mm long, surface above glabrous, surface below sparsely pubescent at base to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 8–17 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers dimorphic, on same individuals (plant monoecious). Flower buds conical to pyramidal. Male inflorescences borne on trunk mainly at ground level, sometimes up to 1 m, composed of 1 to 3 flowers, clumped with other male or female inflorescences. Peduncle inconspicuous. Flower pedicel 95–200 mm long, 1–2 mm in diameter, sparsely pubescent. Bracts 1 to 3 from base towards the lower half of the pedicel, upper bract 1–3 mm long, 1–4 mm wide, depressed ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 2–6.5 mm long, 2–10 mm wide, depressed ovate, free to basally fused, pubescent outside, glabrous inside, color unknown. Petals 3, 11–20 mm long, 9–22 mm wide, length:width ratio 0.8–2, broadly ovate to ovate, fused at base over ca. 30% of the petal length, pubescent to glabrate outside, glabrous inside, pinkish outside. Stamens numerous (exact number unknown), ca. 0.5 mm long, ca. 0.5 mm wide, anthers linear, connective prolongation truncate or absent. Female inflorescences borne on trunk mainly at ground level, sometimes up to 1 m, composed of 1 to 3 flowers, clumped with other male or female inflorescences. Flower pedicel 200–430 mm long, 1–2 mm in diameter, sparsely pubescent. Bracts 1 to 3 from base towards the lower half of the pedicel, upper bract 1–3 mm long, 1–4 mm wide, depressed ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 2–6.5 mm long, 2–10 mm wide, depressed ovate, free to basally fused, pubescent outside, glabrous inside, color unknown. Petals 3, 11–20 mm long, 9–22 mm wide, length:width ratio 0.8–2, broadly ovate to ovate, fused at base over ca. 30% of the petal length, pubescent to glabrate outside, glabrous inside, pinkish outside. Carpels 20 to 40, 2–3.5 mm long, 0.6–1.5 mm wide, densely pubescent, free; stigma ca. 0.5 mm long, ca. 1 mm wide. Fruiting pedicel 200–400 mm long, 1–3 mm in diameter, sparsely pubescent to glabrous. Monocarps, 1–5, 50–80 mm long, 12–32 mm wide, length:width ratio 2.5–3.7, cylindrical, with ca. 4 longitudinal ridge, sparsely pubescent to glabrous, reddish, stipitate; stipe 3–4 mm long, 3–4 mm wide, sparsely pubescent to glabrous. Seeds (2) 6–16 per monocarp, uniseriate to biseriate, ca. 13 mm long, ca. 11 mm wide, ellipsoid, with a longitudinal ridge.
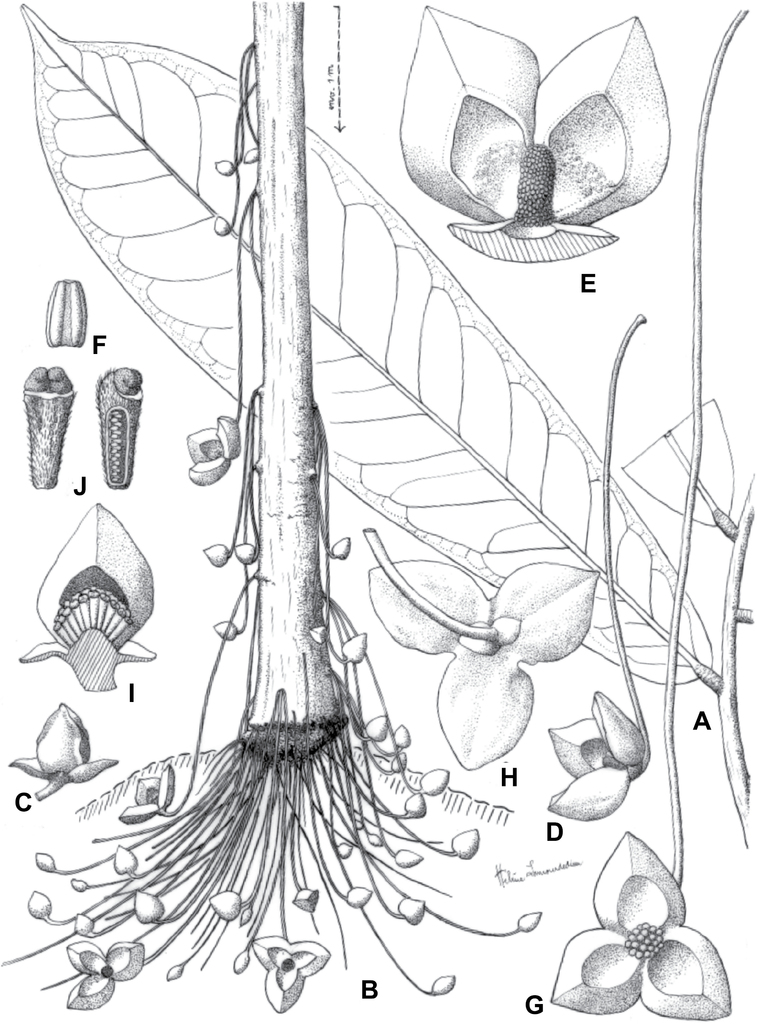
Uvariopsis congolana (De Wild.) Fries A leaves, lower side B trunk with long flowering pedicels C flower bud, side view D male flower, side view, note the long pedicel E detail of male flower, two petals removed, side view F stamen G female flower, top view, note the long pedicel H female flower, bottom view, note the two sepals I longitudinal section of female flower J carpel, front view and detail of ovules A, G from Hallé 3039 B–F, H–J from Hallé 2817. Drawings by Hélène Lamourdedieu, from
Distribution
Element of the Lower Guinean Domain and Congolia Domain of the Guineo-Congolian Region: Democratic Republic of the Congo, Gabon, Republic of the Congo.

Habitat and ecology
Lowland or premontane mature or old secondary rain forest, sometimes in swamp. Altitude: 200–1000 m asl.
Phenology
Flowers collected in March and from August to December. Fruits collected in April, June and December.
Vernacular names
Democratic Republic of the Congo: ‘Asweswe’ in Kimanga (Bokdam 3733), ‘Akobibi’ in Kibila (Ewango 617), ‘Tatakubisa’ (Hart 672).
Notes
Without any flower, this species resemble Up. dioica, Up. guineensis, Up. pedunculosa and Up. solheidii in having oblong to obovate leaves, with acute to rounded base and attenuate to acuminate apex. However, when flowering, Up. congolana differs from all the other Uvariopsis species with flowers with 2 sepals and 3 petals (vs. 2 sepals and 4 petals in other Uvariopsis species). Its female flower pedicels and fruiting pedicels are longer comprised between 20 and 40 cm, which is longer than in the other Uvariopsis species (except Up. pedunculosa that has pedicels that can reach 33 cm). Up. congolana differs from Up. pedunculosa in having less 20 to 40 carpels (vs. 50–140 in Up. pedunculosa), and monocarps marked with longitudinal ridges (vs. monocarps verrucose in Up. pedunculosa).
Conservation status
This species has been previously assessed as Least Concern LC (
Additional specimens examined
Democratic Republic of the Congo – Bandundu • A. Flamigni 10273 (K, M); Mushie, Bolobo; 2°39'03.16'S, 16°21'04.32'E; Mar. 1951 • R. Devred 2881 (K), Kwango, Kiyaka; 5°19'S, 18°56'E; 13 Mar. 1956 – Equateur • C.M. Evrard 5027 (K); Bolomba, Djoa; 0°20'53.88'N, 19°13'44.04'E; alt. 327 m; 15 Oct. 1958 – Orientale • C. Domis 3345 (K); Isangi, Yangambi; 0°46'N, 24°27'E; alt. 470 m; 09 Jan. 1952 • J. Bokdam 3733 (KIS, MO, WAG); Banalia, 20 km along road from Kisangani to Bengamisa; 0°42'N, 25°12'E; 12 Dec. 1972 • J.L.P. Louis 5762 (B, K); Isangi, Yangambi, Isolowe réserve flore; 0°46'N, 24°30'E; 12 Aug. 1937 • T.B. Hart 672 (MO); Mambasa, Epulu; 1°25'N, 28°35'E; alt. 750 m; 14 Aug. 1986 – Unknown major area • C.E.N. Ewango 617 (WAG), Epulu. Zone de Mambasa, Ituri Forest. Afarama; 1°33'N, 28°32'E; alt. 800 m. Gabon – Ogooué-Ivindo • A. Moungazi 234 (P), Bélinga; 1°08'27.49'N, 13°12'03.12'E; alt. 850 m • N. Hallé 2817 (K, P, P, U, WAG), Bélinga; 1°05'N, 13°08'E; alt. 850 m; 27 Oct. 1964 • N. Hallé 3039 (P), Bélinga; 1°05'N, 13°08'E; 06 Nov. 1964 • N. Hallé; A. Le Thomas 64 (K, P), Bélinga, mines de fer, route de Massaka; 1°03'N, 13°12'E; 17 Jul. 1966 – Ogooué-Maritime • J.C. Arends 696 (LBV, MO, P, WAG), Doudou Mountains; 2°15'S, 10°20'E; alt. 670 m; 08 Dec. 1984 • M.S.M. Sosef 1161 (LBV, MO, P, WAG), Monts Doudou, à ± 40 km au Nord-Ouest de Doussala, autour du campement II; 2°13'S, 10°24'E; alt. 425 m; 08 Apr. 2000 – Woleu-Ntem • M.E. Leal 2335 (MO, WAG), Crystal Mountains National Park. Mt. Mekie; 0°26'16'N, 11°13'38'E; alt. 951 m; 20 Nov. 2008. Republic of the Congo – Lékoumou • P. Sita 3209 (P), District de Zanaga, forêt de Mouoni; 2°51'S, 13°50'E; 08 Dec. 1971 – Plateaux • P. Sita 3351 (WAG), région de N'Gouala (Fort-Soufflay); 2°01'S, 14°54'E; 05 Jun. 1972.
Uvariopsis dicaprio
Type
Cameroon – Littoral • L. MacKinnon 51 (holotype: K! (K001381842); isotypes MO, YA); Yabassi, Ebo Forest, Dicam Trail 2000 m from Bekop camp; 4°20'44'N, 10°24'33'E; alt. 849 m; 25 Mar. 2008.
Description
Tree 3–4 m tall, D.B.H 1.8–2.5 cm; young branches glabrous, old branches glabrous. Petiole 4–5 mm long, 1.9–2.1 mm wide, glabrous. Leaf lamina 177–230 mm long, 64–79 mm wide, length:width ratio 2.7–3, obovate, base acute, minutely cordate, apex acuminate, acumen 0.5–1.3 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 5–9 pairs, brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual. Flower buds narrowly ovoid to pyramidal. Male inflorescences borne on trunk from base towards 2.5–3 m, composed of 3 to 7 flowers. Peduncle ca. 2 mm long, ca. 2 mm in diameter, glabrous, bearing radiating 1-flowered partial peduncles 0.5–2 mm long, 0.9–1.2 mm wide. Flower pedicel 18–25 mm long, ca. 1 mm in diameter, glabrous. Bracts 0 to 2, upper bract ca. 1 mm long, ca. 1 mm wide, ovate, sparsely pubescent outside, glabrous inside. Sepals 2, 1–1.5 mm long, 2.1–2.5 mm wide, depressed ovate, glabrous outside, glabrous inside, green to pale brown. Petals 4, 14–16 mm long, 5.5–9 mm wide, length:width ratio 1.7–2.6, ovate to oblong, free, sparsely pubescent outside, glabrous inside, light yellow-green (black when dry) outside, light yellow-green with dark red base inside. Stamens ca. 0.5 mm long, ca. 0.1 mm wide, anthers linear, connective prolongation truncate. Female inflorescences unknown. Fruits unknown.

Uvariopsis dicaprio Cheek & Gosline A inforescences on the trunk B young branch with leaves C inflorescence, side view D detail of male flower, two petals removed E detail of petal outer surface G detail of petiole and base of leaf. A–G from MacKinnon 51 (type). Drawings by Meg Griffiths, from
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region. Known from only one location in Cameroon: the Ebo Forest Reserve.
Habitat and ecology
Submontane mature rain forests. Altitude ca. 280–1000 m asl.
Phenology
Flowers collected in March.
Notes
Up. dicaprio resembles Up. solheidii in having obovate leaves with acute to minutely cordate leaf base and acuminate apex. It differs from Up. solheidii in having young branches and petioles glabrous (vs. tomentose), greater petals (14–16 mm long and 5.5–9 mm wide, vs. 5–10 mm long and 2.5–3.5 mm wide) and petals light yellow-green outside (vs. brown to light red outside). Up. dicaprio also differs from all the other Uvariopsis species in having and inflorescence peduncle ca. 2 mm long and 2 mm wide (vs. inconspicuous).

Preliminary conservation status
This species is known from a single specimen collected in the Ebo National Park, and has previously been preliminarily assessed as Critically Endangered, CR B1+2ab(iii), D (
Uvariopsis dioica
≡ Tetrastemma dioicum
Diels, Bot. Jahrb. Syst. 38(3): 241 (1906). Type. Cameroon – Littoral • H.J.P. Winkler 909 (lectotype: B! (B 10 0153121), lectotype designated in
= Tetrastemma sessiliflorum Mildbr. & Diels syn. nov., Bot. Jahrb. Syst. 53(3–5): 440 (1915); ≡ Uvariopsis sessiliflora (Mildbr. & Diels) Robyns & Ghesq., Ann. Soc. Sci. Bruxelles, Ser. B liii. 322 (1933). Type. Cameroon – East Region • G.W.J. Mildbraed 5239 (holotype: B! (B 10 0153123); isotypes: BR! (BR0000008824233), HBG! (HBG502485)); Haut-Nyong, Lomié; 3°09'N, 13°38'E; 1911.
Description
Tree to shrub 3.6–20 m tall, D.B.H 8–40 cm; young branches pubescent to glabrous, old branches glabrous. Petiole 1.9–5 mm long, 1–2.5 mm wide, pubescent to glabrous. Leaf lamina 111–245 mm long, 38–92 mm wide, length:width ratio 2.5–3.7, elliptic to obovate, base acute to rounded, apex acuminate, acumen 2–30 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 8–14 pairs, brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers dimorphic, on same individuals (plant monoecious). Flower buds ovoid to conical, rarely globose. Male inflorescences borne in clumps, on thickenings of the trunk, mainly between the base and the lower 3 m of the trunk (but up to ca. 5 m on large specimens), composed of 5 to 20 flowers. Peduncle inconspicuous. Flower pedicel (4) 9–55 mm long, 1–1.5 mm in diameter, pubescent to glabrate. Bracts 1 at base and 1 towards the lower half of the pedicel, upper bract ca. 1 mm long, ca. 1 mm wide, broadly ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 1.5–3 mm long, 2.5–5 mm wide, circular to broadly ovate, sparsely pubescent outside, glabrous inside, purplish brown. Petals 4, 6–17 mm long, 3.5–9.5 mm wide, length:width ratio 1.6–2.5, ovate to elliptic, free, valvate, pubescent to glabrous outside, glabrous inside, red to brownish red outside, yellowish cream to reddish at base inside. Stamens ca. 300, 0.2–0.5 mm long, 0.1–0.5 mm wide, anthers linear, connective prolongation truncate. Female inflorescences fewer in number than the male inflorescences, borne in clumps, on thickenings of the trunk, between the base and the lower 25–50 (150 ?) cm of the trunk, composed of 1 to 10 flowers. Flower pedicel (0) 20–77 (128) mm long, 1–2.5 mm in diameter, pubescent to glabrate. Bracts 1 to 3 at base and 1 to 2 towards the lower half of the pedicel, upper bract 1–3 mm long, 1–3 mm wide, broadly ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 2–5 mm long, 4–11 mm wide, circular to broadly ovate, fused at base, sparsely pubescent outside, glabrous inside, purplish brown. Petals 4, 10–25 mm long, 6–17 mm wide, length:width ratio 1.2–2.5, ovate to elliptic, free, valvate (rarely fused at base over 1–3 mm), pubescent to glabrous outside, glabrous inside, red to brownish red outside, yellowish cream to reddish at base inside. Carpels 100 to 280, 1.5–3.5 mm long, 0.7–1.5 mm wide, pubescent, free; stigma 0–0.6 mm long, 0.5–1 mm wide, coiled. Fruiting pedicel 33–97 mm long, 1–3.5 mm in diameter, sparsely pubescent to glabrous. Monocarps, 1–5, 17–85 mm long, 10–50 mm wide, length:width ratio 1.2–2.1, ellipsoid to cylindrical, smooth, not ridged, sparsely pubescent to glabrous, pale brownish grey to yellow to red, sessile to shortly stipitate; stipe up to 3 mm long, 4–5 mm wide, glabrate. Seeds 5–8 per monocarp, biseriate, 15–22 mm long, ca. 15 mm wide, ellipsoid, in a translucid slimy pulp.

Uvariopsis dioica (Diels) Robyns & Ghesq. Fries A trunk with infrutescences B leaves, upper side C leaves, lower side D clump of inflorescences on the trunk E detail of semi-open female flower, side view F fly on petal of femal flower G male flower, top view H detail of female flower, top view (left) and bottom view (right) I transversal cut of fruit J detail of fruit, side view K fruits mature (right) and unripe (left), top view. A, I–K Couvreur 654 B, C Lachenaud 2064 D, G Stevart 4792A E, F, H Bidault 1558. Photos A, I–K Thomas Couvreur B, C Olivier Lachenaud (CC BY-NC-ND 3.0) D, G Tariq Stévart (CC BY-NC-ND 3.0) E, F, H Ehoarn Bidault (CC BY-NC-ND 3.0).
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Cameroon, Equatorial Guinea (Bioko island), Gabon, Nigeria, Republic of the Congo.
Habitat and ecology
Lowland mature or old secondary rain forests. Altitude: 10–980 m asl.
Phenology
Flowers collected from January to June and from October to December. Fruits collected from January to July and from October to November.
Notes
Up. dioica resembles Up. guineensis, Up. pedunculosa and Up. solheidii in having elliptic to obovate leaves, with acute to rounded base and attenuate to acuminate apex. It differs from these species in having flowers borne in clumps of 5 to 20 flowers on thickenings of the trunk, lower than 3 m high (vs. inflorescences of 1 or 2 flowers). Up. dioica has 100 to 240 carpels, whereas all the other Uvariopsis species have less than 100 carpels. Only Up. korupensis and Up. pedunculosa can have up to 120 and 140 carpels, respectively, but Up. dioica has smaller leaves than Up. korupensis (11–25 cm long vs. 28–62 cm long) and smaller flower pedicels than Up. pedunculosa (20–77 mm long vs. 80–325 mm). Here we make Ud. sessiliflora synonym of Ud. dioica based on our phylogenetic analyses (Fig.

Preliminary conservation status
The EOO of this species is estimated at 397,039 km2 and its AOO at 128 km2. Based solely on AOO, it would qualify for Endangered EN under the B2 criterion, but no other subcriterion (a, b or c) is met. Following IUCN criterion B, it is thus assigned a preliminary conservation status of Least Concern LC.
Additional specimens examined
Cameroon – Central Region • G.W.J. Mildbraed 8260 (K), Übergangs – und Kampfgebiet gegen die Savanne an der Nord-grenze der Hylaea südlich des Sanaga zwischen Jaunde und Dengdeng unweit der vereinigung von Lom (Sanaga) und Djerem. Etwa 115 km N O Jaunde; 4°33'04.96'N, 12°16'43.94'E; Mar. 1914 • H. Jacques-Félix 2493 (P), Ndiki; 4°46'N, 10°50'E; Nov. 1938 • P.R.J. Bamps 1458 (P), Résèrve forestière de Makak; 3°29'N, 11°01'E; 14 Dec. 1967 • R.G. Letouzey 12290 (K, P), Mambe près de Boga, 30 km N. Eseka. (feuille IGN 1/200 000 Edea); 3°53'58.29'N, 10°46'58.4'E; 08 Dec. 1973 • R.G. Letouzey 9541 (MO, P, WAG, YA), Versant septentrional des Monts Mfiki au Sud de Ndo, 25 km NNE d'Esse; 4°19'N, 11°58'E; alt. 983 m; 09 Nov. 1969 – Littoral • R.G. Letouzey 12580 (P), Sude de Ngola (8 km Est de l'embouchure de la Sanaga); 3°33'55'N, 9°42'46'E; 05 Jan. 1974 • T.L.P. Couvreur 654 (WAG, YA), Mambe Massif, above Boga village, 100 km along road from Yaoundé to Edea; 3°54'35.85'N, 10°46'25.97'E; alt. 686 m; 19 Jun. 2014 • T.L.P. Couvreur 659 (WAG, YA), Mambe Massif, above Boga village, 100 km along road from Yaoundé to Edea; 3°54'26.07'N, 10°46'21.27'E; alt. 654 m; 20 Jun. 2014 – South Region • J.J.F.E. de Wilde 7754A (K, P, WAG), Roughly between the village of N'Kolandom and Station du Cacaoyer de N'Koemvone; 2°48'N, 11°09'E; 27 Nov. 1974 • J.J.F.E. de Wilde 8270A (BR, MO, P, PRE, WAG), Station du Cacaoyer de N'koemvone, S. of Ebolowa, 14 km on the road to Ambam. On bank of the Seng river; 2°49'N, 11°08'E; alt. 500 m; 05 Jun. 1975 • J.J.F.E. de Wilde 8709 (BR, MO, P, PRE, WAG, YA), Station du Cacaoyer de N'koemvone, S. of Ebolowa, 14 km on the road to Ambam. On bank of the Seng River; 2°49'N, 11°08'E; alt. 550 m; 12 Dec. 1975 • R.G. Letouzey 4219 (K, P), Nkomo, près Ngoassé au S; de la rivière Lobo – feuille IGN 1/200000 Akonolinga; 3°14'N, 12°04'E; 13 Feb. 1962 • R.G. Letouzey 4230 (P), Nkomo, près Ngoassé au S; de la rivière Lobo – feuille IGN 1/200000 Akonolinga; 3°14'N, 12°04'E; 14 Feb. 1962 • R.G. Letouzey 9934 (K, P), près de la rivière Mboro, près Mevous, 50 km SE d'Ebolowa sur piste d'Evindissi; 2°36'N, 11°28'E; 30 Jan. 1970 – South-West Region • G.W.J. Mildbraed 10647 (K), Bibundi-Pflanzung, Westlich des Kamerunberges; 4°12'N, 8°59'E; Nov. 1928 • J.F. Villiers 2429 (P), Pente SW Mt Cameroun, NE Bakingili, 20 km WNW Limbé; 4°05'22.16'N, 9°05'21.91'E; alt. 800 m; 09 Dec. 1984 • J.J. Wieringa 2029 (BR, E, FHO, K, MO, SCA, WAG); Fako, Limbe, W of Njonji Lake; 4°08'N, 9°01'E; alt. 150 m; 27 Jan. 1994 • M.R. Cheek 5482 (K, SCA, YA), Low altitude forest above oil palm plantation. Reached after c. 40 minutes walk N then E from Njonji. Hunters path to ‘Lake Njonji'; 4°08'N, 9°01'E; alt. 300 m; 18 Nov. 1993 • M.R. Cheek 5501 (K, MO, SCA, YA), Low altitude forest above oil palm plantation. Reached after c. 40 minutes walk N then E from Njonji. Hunters path to ‘Lake Njonji'; 4°08'N, 9°00'E; alt. 5 m; 19 Nov. 1993. Equatorial Guinea – Bioko Sur • W.R.Q. Luke 11970 (EA, MA), Moaba – Moka Trail Pt 139 – Pt 138; 3°17'26.92'N, 8°38'29.83'E; alt. 650 m; 16 Mar. 2007 • W.R.Q. Luke 13050 (MA), Ureka pt 330 to Moraka pt 339; 3°15'19.08'N, 8°35'04.56'E; alt. 141 m • W.R.Q. Luke 13113 (EA, MA, MO), Badja E Trail pt 338; 3°18'04.68'N, 8°31'01.2'E; alt. 700 m; 21 Jan. 2009. Gabon – Moyen-Ogooué • E. Bidault 1558 (BR, BRLU, LBV, MO, P, WAG), Est du lac Azingo, à 30 km au Nord-Ouest de Lambaréné; 0°30'32.42'N, 10°05'08.01'E; alt. 48 m; 08 Jun. 2014 • O.L.S. Lachenaud 2064 (MO), piste du lac Azingo, ± 10 km au NE du lac et 30 km au NW de Lambréné; 0°28'41'N, 10°01'59'E; alt. 20 m; 25 Oct. 2014 • T.L.P. Couvreur 926 (LBV, WAG, YA), 27 km after Lambaréné, on road to Bifoum (N1), then around 20 km on road to Lake Azingo; 0°29'38.73'N, 10°01'55.24'E; alt. 23 m; 24 Nov. 2015 • T.O.B.E.B. Stévart 4792A (P, WAG), Okala, North of Libreville. Terrain de Montigny à Okala (Libreville); 0°29'N, 9°25'E; alt. 10 m; 12 Feb. 2014 – Ogooué-Ivindo • A. Hladik 1472 (BRUNOY, P, P, US), Ipassa, au croisement des layons J et VII; 0°30'N, 12°48'E; alt. 490 m; 29 Oct. 1971 • J.M. Reitsma 3507 (LBV, MO, WAG), Primary rain forest, near village Ekobakoba, 50 km SE of Makokou; inventory; 0°23'N, 13°06'E; 21 May. 1987 • L.J. Dorr 4248 (LBV, MO), Station I.R.E.T. (M'Passa Field Station). 10 km S de Makokou sur la riviere Ivindo; 0°30'N, 12°45'E; alt. 500 m; 13 May. 1985 • L.J. Dorr 4275 (LBV, MO, P), Station I.R.E.T. (M'Passa Field Station), 10 km S de Makokou sur la riviere Ivindo; 0°30'N, 12°45'E; alt. 500 m; 14 May. 1985 • M.S.M. Sosef 2210 (BR, LBV, MO, WAG), Ipassa Reserve, IRET Research Station, SW of Makokou; 0°31'N, 12°48'E; alt. 350 m; 04 Nov. 2005 – Woleu-Ntem • A.M. Louis 4098 (BR, LBV, MO, WAG), 25 km NE of Bitam, Nsimy; 2°08'N, 11°22'E; 02 May. 1995. Nigeria – Edo State • A.F. Ross 2/6 (K); Benin, Sapoba Forest Reserve; 6°06'N, 5°53'E; 1934 • B.O. Daramola FHI72315 (K, MO), Iyekoriowon. Forest Reserve. In P.S.P. 82 Ugo; 6°05'19.05'N, 6°00'04.46'E; 04 Oct. 1973 • R.D. Meikle 628 (K, P), Usonigbe Forest Reserve near Sapoba; 5°56'17.52'N, 5°53'20'E; 16 Nov. 1944 • R.W.J. Keay FHI28066 (K, K, P); Benin, Sapoba Forest Reserve, Compte.9,S.S.I; 6°06'N, 5°53'E; 03 Nov. 1950 – Kogi State • M.G. Latilo FHI47768 (K); Kabba, Southern Adoru Forest Reserve, Igala, bank of river Owe near Adoru; 6°55'38.49'N, 7°05'38.55'E; 01 Jul. 1963. Republic of the Congo – Pool • J. Koechlin 4001 (P), Bangou, Forêt de; 3°54'S, 14°27'E; Dec. 1956 • P. Sita 3264 (P), Forêt de Bangou, Est de M'Passa; 4°29'34'S, 14°45'28'E; 18 Dec. 1971.
Uvariopsis etugeana
Type
Cameroon – North-West Region • R.G. Letouzey 13414 (holotype: P! (P01982826)), Rive droite de la Metchum, près Obang (18 km S of Wum); 6°15'N, 10°01'59.99"E; alt. 600 m; 03 Dec. 1974.
Description
Tree 3–6 m tall, D.B.H unknown; young branches slightly pubescent, old branches glabrous. Leaves with minute pelucid punctuations. Petiole 3.5–4 mm long, 2.5–3.5 mm wide, glabrous. Leaf lamina 190–235 mm long, 70–85 mm wide, length:width ratio ca. 2.7, elliptic, coriaceous, base acute to slightly decurrent, apex attenuate to acuminate, acumen ca. 10 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 8–10 pairs, brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers dimorphic, on same individuals (plant monoecious). Flower buds ovoid to pyramidal. Male inflorescences borne on trunk, composed of 1 to 2 flowers. Peduncle inconspicuous. Flower pedicel 8–10 mm long, 1–1.5 mm in diameter, glabrate to glabrous. Bracts 1 at base and 1 towards the middle or lower half of the pedicel, upper bract 1–1.5 mm long, 1–2.5 mm wide, broadly ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, ca. 2 mm long, ca. 4 mm wide, broadly ovate, basally fused, sparsely pubescent to glabrous outside, glabrous inside, color unknown. Petals 4, 6–11.5 mm long, 3.5–7 mm wide, length:width ratio 1.6–2.5, elliptic to ovate, sparsely pubescent to glabrous outside, glabrous inside, color unknown. Stamens numerous (exact number unknown), 0.5–1 mm long, ca. 0.2 mm wide, anthers linear, connective prolongation truncate. Female inflorescences borne on trunk, composed of 1 to 2 flowers. Flower pedicel ca. 4 mm long, ca. 2 mm in diameter, glabrate to glabrous. Bracts 1 at base and 1 towards the middle or lower half of the pedicel, upper bract ca. 1.5 mm long, ca. 2.5 mm wide, broadly ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 1–1.5 mm long, 2–3.5 mm wide, broadly ovate, basally fused, sparsely pubescent to glabrous outside, glabrous inside, color unknown. Petals 4, ca. 14 mm long, ca. 8 mm wide, length:width ratio 1.6–2.5, elliptic, glabrous outside, glabrous inside, color unknown. Stamens ca. 5, ca. 1 mm long, sterile, seen on one female flower. Carpels ca. 20, ca. 3 mm long, ca. 1 mm wide, glabrate at base to glabrous, free; stigma coiled. Fruits unknown.

Uvariopsis etugeana Dagallier & Couvreur A branch with leaves B flower, semi-bottom view C inflorescence with two flowers D female flower, transversal cut, three petals removed E male flower, transversal cut of the receptacle. A–C Letouzey 13414 D, E Thomas 4544. Photos Léo-Paul Dagallier.
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region. Known from only two localities in the North-West and South-West Regions in Cameron.

Habitat and ecology
Lowland mature rain forests or semi-deciduous forests. 170–600 m asl.
Phenology
Flowers collected in March and December.
Notes
Up. etugeana resembles Up. pedunculosa in the shape of the leaves, and Up. solheidii in the shape of its flowers. However, Up. etugeana has a short flowering pedicel (4–10 mm vs. 14–325 mm in Up. pedunculosa and 9–198 mm in Up. solheidii) and petals which are glabrous outside (vs. pubescent in both Up. pedunculosa and Up. solheidii).
Preliminary conservation status
This species is known from only two specimens from two different locations in western Cameroon. Its EOO is estimated less than 1,000 km2 and its AOO is estimated at 8 km2. The two specimens were collected more than 30 years ago, and only one occurs in a protected area, in the Takamanda National Park. This species is thus likely to have undergone, or to undergo in the future, a decline in its AOO. Following IUCN criterion B, it is thus assigned a preliminary conservation status of Endangered EN B1ab(i,ii)+2ab(i,ii).
Additional specimens examined
Cameroon – South-West Region • D.W. Thomas 4544 (MO), Takamanda Forest Reserve; 6°14'N, 9°19'E; alt. 170 m; 21 Mar. 1985.
Uvariopsis guineensis
≡ Uvaria spectabilis A.Chev., Explor. Bot. Afrique Occ. Franc. i. 7 (1920). Type. Ivory Coast – Danané • A.J.B. Chevalier 21305 (holotype: P! (P00362614), sheet here designated; isotype: P! (P00362612)), Haut Cavally, Pays des Byolas, entre Danané et Goutokouma; 7°16'N, 8°09'W; 25 Apr. 1909.
= Uvariopsis globiflora Keay; syn. nov. concerning Uvariopsis guineensis var. globiflora (see details under this variety).
Description
Tree 2.5–9 m tall, D.B.H 4–12 cm; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 1.5–5 mm long, 1.5–3.5 mm wide, sparsely pubescent to glabrous. Leaf lamina 110–310 mm long, 45–108 mm wide, length:width ratio 2.3–4.2, elliptic to oblong to obovate, coriaceous, base acute to rounded, apex attenuate to acuminate, acumen 9–30 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, sparsely pubescent to glabrous below; secondary veins 7–14 pairs, brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers dimorphic, on same individuals (plant monoecious). Flower buds globose to oblate. Male inflorescences borne on trunk, axillary or terminal, composed of 1 flower. Peduncle inconspicuous. Flower pedicel 5–16 mm long, 1–2 mm in diameter, sparsely pubescent. Bracts 1 at base and 1 towards the middle or lower half of the pedicel, upper bract 1–3.5 mm long, 1–3.5 mm wide, broadly ovate, adpressed, semi-clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 1–3 mm long, 2–7 mm wide, broadly ovate, free to basally fused, pubescent to sparsely pubescent outside, glabrous inside, yellowish green. Petals 4, 7–17 mm long, 5–17 mm wide, length:width ratio 0.9–1.4, broadly ovate, free to fused at base over 30–50% of their length, pubescent to sparsely pubescent outside, glabrous inside, pale greenish to cream outside, greenish cream to cream with purplish base inside. Stamens number unknown, 0.3–1 mm long, 0.2–0.3 mm wide, anthers linear, connective prolongation truncate. Female inflorescences borne on trunk, axillary or terminal, composed of 1 flower. Flower pedicel 10–65 mm long, 1–2 mm in diameter, sparsely pubescent. Bracts 1 to 3 at base and 1 towards the middle or lower half of the pedicel, upper bract 1–3.5 mm long, 1–3.5 mm wide, broadly ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 1–6 mm long, 2–10 mm wide, broadly ovate, free to basally fused, sparsely pubescent to pubescent outside, glabrous inside, yellowish green. Petals 4, 9–24 mm long, 7–22 mm wide, length:width ratio 0.9–1.4, broadly ovate, free to fused at base over 30–50% of their length, pubescent to sparsely pubescent outside, glabrous inside, pale greenish to cream outside, greenish cream to cream with purplish base inside. Carpels 10 to 50, 2–5 mm long, 0.6–1.1 mm wide, pubescent to velutinous, free; stigma ca. 0.5 mm long, 0.5–0.8 mm wide, coiled. Fruiting pedicel 17–69 mm long, 1.5–4.5 mm in diameter, sparsely pubescent to glabrous. Monocarps, 2–10, 20–70 (90) mm long, 12–25 mm wide, length:width ratio 1.7–3.3 (measures taken from both dried fruits and their associated specimen label, note that monocarps seem to sink when drying), cylindrical, smooth, straight to slightly curved, with a longitudinal ridge, sparsely pubescent to glabrous, dull olive green to red to brown, sessile to shortly stipitate; stipe up to 3 mm long, 2–6 mm wide, pubescent to glabrous. Seeds 2–22 per monocarp, biseriate, 12–17 mm long, 10–13 mm wide, in a yellow pulp with strong aniseed scent.

Uvariopsis guineensis Keay A leaf, lower side B leaf, upper side C fruits on trunk D old female flower, petals fallen, side view E flower, semi-bottom view F detail of female flower, petals removed, side view G male flower, top view H monocarp, longitudinal cut. A, B, D–F Bidault 4798 C, H Bidault 632 G Koivogui 250. Photos A–G Ehoarn Bidault (CC BY-NC-ND 3.0)
Distribution
Endemic to Upper Guinean Domain of the Guineo-Congolian Region: Ghana, Guinea, Ivory Coast, Liberia, Sierra Leone.
Habitat and ecology
Lowland mature or secondary rain forests. Altitude: 100–950 m asl.
Phenology
Flowers collected from March to May and from September to December. Fruits collected from March to July and in September.
Notes
Up. guineensis resembles Up. dioica, Up. pedunculosa, and Up. solheidii in having elliptic to oblong to obovate leaves, with acute to rounded base and attenuate to acuminate apex. Young branches and petioles of Up. guineensis are sparsely pubescent to glabrous (vs. pubescent to glabrous in Up. dioica and Up. pedunculosa, and tomentose to shortly tomentose in Up. solhdeidii). Up. guineensis has less than 50 carpels (vs. more than 50 in Up. dioica and Up. pedunculosa), and its monocarps are smooth with a single longitudinal ridge (vs. with longitudinal and transversal ridges in Up. solheidii and verrucose in Up. pedunculosa). Here we make the names Ud. guineensis and Ud. globiflora synonyms. They have traditionally been discriminated based on the position of their flowers and the fusion of the petals, with Ud. globiflora having axillary or terminal flowers and free petals and Ud. guineensis having flowers borne on trunk (cauliflory) and fused petals (

Conservation status
This species is widespread in West Africa, from Sierra Leone to Ghana. It has been assessed as Least Concern LC (
Uvariopsis guineensis var. globiflora , stat. nov.
≡ Uvariopsis globiflora Keay syn. nov., Kew Bull. 7(2): 152 (1952). Type. Ghana – Eastern Region – C. Vigne FH1877 (holotype: K! (K000041224), sheet here designated; isotypes: K! (K000041225, K000041226)), Amentia; 6°13'N, 1°10'W; alt. 160 m; Mar. 1930.
Description
Male inflorescences axillary or terminal (more rarely borne on trunk). Petals 7–10 mm long, 5–9 mm wide, length:width ratio 1.1–1.4, broadly ovate, free. Female inflorescences axillary or terminal (rarely borne on trunk). Sepals 1–3 mm long, 2–4 mm wide. Petals 10–22 mm long, 9–17 mm wide, length:width ratio 1.1–1.4, broadly ovate, free. Female flowers: carpels 10 to 20, 2–4 mm long, 1–1.1 mm wide.
Distribution
Endemic to Upper Guinean Domain of the Guineo-Congolian Region: Ghana.
Habitat and ecology
Lowland mature or secondary rain forests. Altitude: 160–800 m asl.
Notes
Up. guineensis var. globiflora differs from the type variety in having free petals (vs. fused at base over 30–50% of their length) and fewer carpels (10–20 carpels vs. 20–50 carpels). The male flowers have also smaller petals (7–10 mm long and 5–9 mm wide vs. 8–17 mm long and 9–17 mm wide).
Additional specimens examined
Ghana – Ashanti Region • J.E. Andoh 5096 (P), Bobiri Forest Reserve; 6°40'N, 1°20'W; Jan. 1948 • J.E. Andoh FH4246 (K, P), South Fomangsu Reserve; 6°36'N, 0°57'E; Oct. 1936 – Brong-Ahafo Region • A.A. Enti FE1220 (K, MO), Asukese F.R; 7°09'07'N, 2°31'07'W; 02 Mar. 1973 – Eastern Region • A.A. Enti GC37451 (K), Atewa Range F.R; 6°09'N, 0°36'E; 14 Dec. 1967 • A.A. Enti R698 (BR, MO), Aiyaola Forest Reserve, Kade; 6°09'N, 0°57'E; 03 May. 1972 • C.C.H. Jongkind 1809 (MA, MO, WAG), Atewa Range Forest Reserve, along Old Geological Survey road; 6°14'06'N, 0°33'E; alt. 400 m; 26 Oct. 1994 • D.K. Harder 3325 (MO, WAG), Asiakwa District: Sagyimase Village; Atewa Range (Forest Reserve); between 6–8 km NW of intersection of Accra-Kumasi Road at Sagyimase along forest access road; 6°13'48'N, 0°32'42'E; alt. 810 m; 04 Jul. 1995 • J.K. Morton A4247 (K, WAG), Kade Agric. Res. Station; 6°08'N, 0°55'E; 04 May. 1961 – Western Region • A. Foggie 4456 (K), Bia National Park, Bia F. Resrve; 6°29'55'N, 3°05'24"W; Apr. 1937.
Uvariopsis guineensis var. guineensis
Description
Male inflorescences borne on trunk, axillary or terminal. Petals 8–17 mm long, 9–17 mm wide, length:width ratio 0.9–1.2, broadly ovate, fused over 30–50 % of their length. Female inflorescences borne on trunk, axillary or terminal. Sepals 3–6 mm long, 5–10 mm wide. Petals 9–24 mm long, 7–22 mm wide, length:width ratio 0.9–1.3, broadly ovate, fused at base over 30–50% of their length. Carpels 20–50, 2.1–5 mm long, 0.6–1 mm wide.
Distribution
Endemic to Upper Guinean Domain of the Guineo-Congolian Region: Ghana, Guinea, Ivory Coast, Liberia, Sierra Leone.
Habitat and ecology
Endemic to Upper Guinean Domain of the Guineo-Congolian Region: Lowland mature or secondary rain forests. Altitude: 100–950 m asl.
Additional specimens examined
Ghana – Western Region • H.H. Schmidt 2070 (MO), Bia National Park and Production Reserve. From the town of Bankasa, PArk Guard Camp 13, along foot trail to Park Guard Camp 11; 6°32'40'N, 3°08'20'W; alt. 180 m; 05 Mar. 1996. Guinea – Nzérékoré • A.J.B. Chevalier 20797 (K, P); Macenta, Pays des Koniankéi, Fassakoïdou; 8°40'N, 9°17'W; 24 Feb. 1909 • C.C.H. Jongkind 10660 (WAG), Forêt Classée de Mt Yonon, not far from the Diane River; 7°58'58.8'N, 9°07'16.8'W; alt. 480 m; 07 May. 2011 • C.C.H. Jongkind 10774 (BR, MO, P, WAG), Forêt Classée de Mt Yonon, not far from the Diane River; 7°59'01.8'N, 9°07'17.4'W; alt. 445 m; 12 May. 2011 • C.C.H. Jongkind 11343 (MO, WAG), Nimba mountains. East side of main ridge; 7°38'56.4'N, 8°21'40.8'W; alt. 793 m; 30 Jun. 2012 • C.C.H. Jongkind 7767 (BR, K, P, WAG), Nimba Mountains, banks of Zié River close to reserve boundary; 7°42'58.2'N, 8°21'39.6'W; alt. 540 m; 21 Jun. 2007 • D. Bilivogui 4 (MO, P, WAG); Lola, Guinée Forestière, Nimba Mountains, SMFG iron ore mine concession. Gouan River valley, at its lowest point inside the reserve; 7°42'46'N, 8°23'40'W; alt. 535 m; 28 Sep. 2011 • E. Bidault 4798 (BRLU, MO, P, SERG), Monts Nimba, autour du camp Seringbara 2; 7°37'43.5'N, 8°25'39.72'W; alt. 1020 m; 09 Oct. 2019 • E. Bidault 632 (MO, P); Lola, Nimba Mountain, Guinea, Monts Nimba, site du patrimoine mondial, forêt de Gbié au nord de la grande savane; 7°56'38.37'N, 8°21'38'W; alt. 773 m; 26 Jun. 2012 • J.-G. Adam 3826 (WAG), Nzérékoré. Jomou; 24 Feb. 1949 • Nimba Botanic Team JR1272 (WAG), Nimba Mountains, plot JRFM01, vallée de Gba; 7°40'34.8'N, 8°23'30'W; alt. 947 m; 05 Dec. 2007 • P.K. Haba 107 (K), Béro Mountains, Le sommet du Mont Thon dans les Monts Béro. Proche de la ville de Boola; 8°19'12'N, 8°41'47'W; alt. 1099 m; 03 Dec. 2007 • R.A.A. Schnell 3618 (P), Mt Nimba., Blanmbaya; 7°37'N, 8°25'W; Sep. 1947 • X.M. van der Burgt 1300 (K, WAG); Macenta, Guinée Forestière. Macenta + Beyla préfectures. Simandou Range. South of Pic de Fon, 1.4 km south and 900 m west of Fokou hill; 8°28'55'N, 8°54'05'W; alt. 855 m; 15 Sep. 2008. Ivory Coast – Abengourou • C. Versteegh 619 (U, WAG), Surroundings of Niablé, 30 km east of Abengourou; 6°39'40.63'N, 3°16'11.42'W; 31 Jul. 1969 – Abidjan • J. de Koning 4587 (WAG), Experimental Station ORSTOM, Adiopodoumé. Seedlings, seed source Banco Forest; 5°23'N, 4°03'W; 30 Oct. 1974 • J. de Koning 4905 (WAG), CULTA. Experimental Station ORSTOM, Adiopodoumé. Seedlings, seed source Banco Forest; 5°23'N, 4°03'W; 29 Nov. 1974 • J. de Koning 5170 (MO, WAG), Abidjan, Banco Forest; 5°23'N, 4°03'W; 16 Jan. 1975 • J. de Koning 5504 (BR, MO, WAG), Banco Forest; 5°23'N, 4°03'W; 08 Mar. 1975 – Daloa • L.P.G.A. Nusbaumer 36 (G), F.C. Scio., Pinhou, Lobykro à 5 Km, Bloc 28, Parcelle 141 de la Sodefor; 7°42'N, 6°50'W; 10 Sep. 2001 – Danané • L. Aké Assi 8823 (G), Forêt de Tiapleu 7.4197, -8.2116; 7°25'10.92'N, 8°12'57.6'W; 30 Apr. 1966 • L. Aké Assi 9131 (G), Momi Mont, Foret Classee du, Forêt de Mont Momi; 7°24'N, 8°03'W; 28 Oct. 1966 – Soubré • A.J.B. Chevalier 19224 (P), entre le moyen Sassandra et le moyen Cavally; 5°55'59.75'N, 7°03'46.35'W; 06 Jul. 1907 – Vavoua • F.N. Kouamé 1445 (CSRS, G), F.C. du Haut-Sassandra, Nord. forêt dégradée, relevé FNK14; 7°18'N, 7°01'W; 04 Apr. 1995. Liberia – Bong • D.H. Linder 580 (K), Gbanga; 7°00'N, 9°29'W; 12 Sep. 1926 – Grand Gedeh • C.C.H. Jongkind 6667 (WAG), along Zwedru – Harper road, south of Tiama Town; 5°34'12.6'N, 8°09'16.2'W; alt. 260 m; 07 Jun. 2005 – Lofa • C.C.H. Jongkind 6691 (WAG), North Lorma National Forest; 8°03'N, 9°42'W; 19 Nov. 2005 • C.C.H. Jongkind 9478 (WAG), near Ziggida, rappids in Jèbèh River; 8°02'10.8'N, 9°28'55.8'W; alt. 455 m; 12 Feb. 2010 – Montserrado • J.J.F.E. de Wilde 3811 (MO, WAG), between Bomi hills and Lofa river, c. 15 km N.N.W. of Bomi hills; 6°58'N, 10°52'W; 15 Apr. 1962 – Nimba • J.-G. Adam 20713 (IFAN, K, P, UPS), Nimba; 7°32'N, 8°32'W; alt. 450 m; 23 Jan. 1965 • J.-G. Adam 26362 (MO), Granfield Mt Nimba; 7°34'N, 8°29'W; 17 Oct. 1971 • Nimba Botanic Team NS74 (WAG), Valley between Mt Gangra and Mt Yuelliton, plot SNFR01; 7°33'22.2'N, 8°38'W; alt. 700 m; 14 Jan. 2009 • W.J. Harley 1556 (K), Bilimu; 7°24'N, 8°36'W; 06 Jan. 1948 – Sino • B. Senterre 7066 (MO), Dugbe hummingbird site (ca. 20 km South of Sapo National Park, ca. 50 km East of Greenville), North of Money camp; 5°06'47'N, 8°32'18'W; alt. 107 m; 27 Mar. 2014 • C.C.H. Jongkind 8898 (WAG), Babooni road not too far from main road; 5°31'N, 8°37'36'W; alt. 140 m; 09 Mar. 2009. Sierra Leone – Eastern Province • D. Small 101 (K); Kenema, Dambayei Valley. Kenema, Kambui Hills F.R; 7°48'N, 11°11'W; 25 May. 1951 • J. Momoh 37 (K, SL, WAG), Gola National Park, central block. 600 m N of Malimbe camp; 7°40'07'N, 10°53'22.7'W; alt. 320 m; 24 Oct. 2013 – Northern Province • J.-G. Adam 23345 (MO); Koinadugu, Loma Mountains, Kabala Mt Loma – Kamia; 9°22'37.15'N, 11°19'26.51'W; 26 Jan. 1966 • P. Jaeger 7488 (P); Koinadugu, Loma Mountains, Loma; 9°10'N, 11°07'W; 16 Sep. 1964 • P. Jaeger 7489 (G); Koinadugu, Loma Mountains, W Loma; 9°10'N, 11°07'W; alt. 300 m; 16 Sep. 1964 • P. Jaeger 8041 (G, K, P); Koinadugu, Mt Loma; 9°10'N, 11°07'W; alt. 600 m; 13 Oct. 1964 • P. Jaeger 8756 (K); Koinadugu, Mt Loma; 9°10'N, 11°07'W; alt. 800 m; 29 Dec. 1965 • P. Jaeger 9093 (K, P); Koinadugu, Mt Loma; 9°10'N, 11°07'W; 26 Jan. 1966 • P. Jaeger 9159 (G, K, P); Koinadugu, Mt W Loma; 9°10'N, 11°07'W; alt. 600 m; 29 Jan. 1966 – Southern Province • A.H. Unwin 51 (K); Pujehun, Gola Forest; 7°16'N, 11°18'W; 1909 • D. Small 593 (K, K, P); Pujehun, Gola forest; 7°40'N, 10°55'W; 08 Apr. 1952 • D. Small 655 (K); Pujehun, Gola Forest; 7°16'N, 11°18'W; 1952 • D. Small 676 (K, P); Pujehun, Gola forest, Bagbe Line 20n B.III; 7°16'N, 11°18'W; 19 May. 1952 • D. Small 678 (K); Pujehun, Gola forest, Bagbe Line 20n B.III; 7°16'N, 11°18'W; 15 May. 1952 • H.D. Jordan 2068 (B, K, P); Pujehun, Gola Forest, mile 44 (Tunkia); 7°39'N, 10°54'W; 14 May. 1955.
Uvariopsis korupensis
Type
Cameroon – South-West Region • D. Kenfack 1026 (holotype: YA! (YA0003173); isotypes: MO! (MO-022919), P! (P01817719), WAG! (WAG0358388)); Ndian, Korup National Park, Chimpanzee Camp; 5°04'N, 8°52'E; alt. 160 m; 03 Feb. 1998.
Description
Tree 3–15 m tall, D.B.H 5–14 cm; young branches pubescent to glabrous, old branches glabrous. Petiole 2–7 mm long, 3–6 mm wide, pubescent to glabrous. Leaf lamina 280–615 mm long, 83–165 mm wide, length:width ratio 2.9–4.3, obovate, coriaceous, base rounded to cordate, apex acuminate, acumen 18–32 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 13–26 pairs, brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers dimorphic, on same individuals (plant monoecious, but individuals with only staminate flowers were also observed). Flower buds conical. Male inflorescences borne on thickenings of the trunk, mainly at the base of the trunk, towards up to 3.8 m, composed of 2 to 3 flowers. Peduncle inconspicuous. Flower pedicel 6–70 mm long, 1–2 mm in diameter, pubescent to sparsely pubescent. Bracts 1 to 3 at base and one towards the lower 10% of pedicel, upper bract 1–3.5 mm long, 1.5–5 mm wide, broadly ovate, adpressed, semi-clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 1–5 (7.5) mm long, 2–6.5 mm wide, broadly ovate, basally fused, pubescent outside, glabrous inside, dull yellowish-brown. Petals 4, 10–38 mm long, 4–10 mm wide, length:width ratio 2.5–7, narrowly ovate, fused at base, pubescent outside, glabrous and verrucose inside, cream to brownish outside, cream to pinkish inside. Stamens numerous (exact number unknown), 0.5–0.8 mm long, 0.1–0.3 mm wide, anthers linear, connective prolongation truncate. Female inflorescences borne on thickenings of the trunk, mainly at the base of the trunk, towards up to 3.8 m, composed of 2 to 3 flowers. Flower pedicel 20–50 mm long, 1.5–2.5 mm in diameter, pubescent to sparsely pubescent. Bracts 1 to 3 at base and one towards the lower 10% of pedicel, upper bract 1–3.5 mm long, 1.5–5 mm wide, broadly ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 3–5 mm long, 4–5 mm wide, broadly ovate, basally fused, pubescent outside, glabrous inside, dull tan. Petals 4, 15–35 mm long, 7–12 mm wide, length:width ratio 2.2–5, narrowly ovate, fused at base, pubescent outside, glabrous and verrucose inside, cream to brownish outside, cream to pinkish inside. Carpels 25 to 120, 1.5–2.5 mm long, 0.5–1 mm wide, velutinous, free; stigma ca. 0.6 mm long, 0.8–1.3 mm wide, truncate pyramidal. Fruiting pedicel 45–90 mm long, ca. 3 mm in diameter, glabrous. Monocarps, 5–9, 29–45 mm long, 18–30 mm wide, length:width ratio 1.4–1.9, ellipsoid to cylindrical, smooth, glabrous, clear orangish yellow, sessile to shortly stipitate; stipe up to 5 mm long, ca. 2 mm wide, glabrous. Seeds 8–14 per monocarp, biseriate, 10–22 mm long, 5–14 mm wide, ellipsoid to oblong.

Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Cameroon, Equatorial Guinea, Gabon.
Habitat and ecology
Lowland mature or secondary rain forests. Soil: sandy, rocky, volcanic. Altitude 90–160 m asl.
Phenology
Flowers collected from January to March, in April and from July to November. Fruits collected from February to April and in October.
Notes
This species resembles Up. bakeriana, Up. citrata and Up. submontana in having large obovate leaves (from 28 to 62 cm long), with rounded to cordate bases. Up. korupensis differs from Up. bakeriana and Up. citrata in having longer flower pedicel (6–70 mm vs. less than 8 mm), and differs from Up. bakeriana in having fused and cream petals (vs. free and pinkish red). It is harder to differentiate Up. korupensis and Up. submontana. Up. korupensis has generally larger leaves than Up. submontana (28–62 cm vs. 16–38 cm). The female flowers of Up. korupensis have smaller sepals (3–5 mm long vs. 6–8 mm long in Up. submontana), and generally longer petals (15–35 mm long, vs. 15–17 mm long), which results in petals more than 3 times longer than the sepals (vs. petals less than 3 times longer than the sepals in Up. submontana). Petals of Up. korupensis are cream to brownish outside and cream to pinkish inside whereas petals of Up. submontana are pinkish to dark red inside and outside (Table

Uvariopsis korupensis Gereau & Kenfack A branch with leaves B female flower, side view C female flower, three petals removed, side view D carpel E male flower and male flower bud, side view F male flower, one petal removed, side view G male flower, bottom view H petal of male flower, inner view I fruit, with A single monocarp J longitudinal section of monocarp. A from Kenfack 1146 B–K from fresh material. Scale bars: 1 cm (A–C, E–K); 0.5 cm (D). Drawings by David Kenfack, from “Le genre Uvariopsis (Annonaceae) en Afrique tropicale, avec la description d’une espèce nouvelle du Cameroun” Adansonia 22:1, fig. 1, p. 42 (
Conservation status
This species has been assessed as Endangered EN under criteria B2ab(iii) (
Additional specimens examined
Cameroon – Littoral • D. Kenfack 1620 (MO), Nkam, Yingui. Ekem River bank; 4°32'N, 10°18'E; alt. 500 m; 05 Mar. 2002 – South Region • G.A. Zenker 3971 (BM, E, G, G, K), Bipinde; 3°05'N, 10°25'E; 1911 • G.P. Tchouto Mbatchou BIFAX25 (WAG), Campo Ma'an area, Bifa, in the National Park; 2°40'28'N, 10°16'54'E; alt. 60 m; 13 Oct. 2001 – South-West Region • A.H. Gentry 62456 (MO), Banyong, Batanga area, between Awong and Banyu, ca 15 km W of Manyemen. TRANSECT 3; 5°00'N, 9°10'E; alt. 420 m; 03 May. 1988 • D.W. Thomas 3182 (L, MO, P); Ndian, Cameroon. South-West, forest in the Korup National Park; 5°03'N, 8°48'E; alt. 50 m; 28 Feb. 1984 • D.W. Thomas 631 (K); Ndian, Korup National Park, transect R; 5°06'N, 8°55'50'E; Jul. 1979 • G.P. Tchouto Mbatchou 675 (K, SCA, YA), Bomana forest. Transect OA, Plot OA0X; 4°15'N, 9°01'E; alt. 200 m; 05 Oct. 1993 • J. Nning 284 (K, MO, SCA, YA); Fako, Bakingili, along a hill`slope to drink gari Camp above the 6th plot; 4°05'N, 9°03'E; alt. 480 m; 16 Feb. 1997 • M.R. Cheek 5486 (K, SCA, YA), Low altitude forest above oil palm plantation. Reached after c. 40 minutes walk N then E from Njonji. Hunters path to ‘Lake Njonji'; 4°08'N, 9°01'E; alt. 300 m; 19 Nov. 1993 • M.R. Cheek 8258 (K, YA); Ndian, Korup National Park, Ekundu Kundu, Transect 9, c. 7.5 cm; 5°07'N, 8°50'E; alt. 170 m; 28 Apr. 1996 • M.R. Cheek 8815 (K, YA); Ndian, Korup National Park, path from village of Ekundu Kundu to top of Mt Juahan; 5°09'N, 8°52'E; alt. 550 m; 09 Jan. 1998 • R.E. Gereau 5192 (MO, P, WAG), Ndian Division; NW of Mundemba and W of Fabe Village, less than 1 km NE of confluence of Six Cup Garri Creek with Ndian (Mana) River; 5°03'N, 8°53'E; alt. 90 m; 05 Mar. 1993 • T.L.P. Couvreur 1039 (MPU, WAG, YA), Mount Cameroon National Park, Bakinguili trail, above Bakinguili village; 4°05'47.97'N, 9°03'24.72'E; alt. 563 m; 02 Apr. 2016 • T.L.P. Couvreur 1052 (MPU, WAG, YA), Mount Cameroon National Park, on the Bomona trail, behind Bomona village, 10 km NW from Idenau; 4°17'48.99'N, 9°04'43.07"E; alt. 688 m; 03 Apr. 2016. Equatorial Guinea – Rio Muni, Litoral • C.M. Wilks 1784 (LBV, WAG), Monts de Cristal. 11 km ENE d'Okuamkos; 1°09'N, 10°14'E; 10 Aug. 1988 – Unknown major area • C.M. Wilks 1818 (LBV, P, P, WAG), Monts de Cristal. 15 km d'Okuamkos; 1°10'N, 10°16'E; 23 Aug. 1988. Gabon – Estuaire • A.M. Louis 1863 (BR, K, LBV, MO, WAG), Andem, Kougouleu-Kango ± 10 km, 2 km N; 0°22'N, 9°57'E; 09 Oct. 1985.

Uvariopsis lovettiana
Type
Tanzania – Morogoro • T.L.P. Couvreur 97b (holotype: WAG! (WAG0361229), sheet here designated; isotypes: DSM, MO, WAG! (WAG0361230, WAG0361231)); Kilombero District, E Udzungwa National Park. In forest S of Mwanihana hill. c. 2 km S of last camping site on Mwanihana trail; 7°48'35.36'S, 36°49'26.4'E; alt. 1400 m; 30 Nov. 2006.
Description
Tree 3–30 m tall, D.B.H 3.5–50 cm; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 4–9 mm long, 1.2–1.5 mm wide, sparsely pubescent to glabrous. Leaf lamina 150–280 mm long, 45–72 mm wide, length:width ratio 2.5–4, elliptic, coriaceous, base acute to decurrent, apex acute to acuminate, acumen 5–25 mm long, surface above glabrous, surface below sparsely pubescent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 9–16 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers similar, on same individuals (plant monoecious). Flower buds globose. Male and female inflorescences axillary, composed of 1 flower. Peduncle inconspicuous. Flower pedicel 14–50 mm long, 0.7–3 mm in diameter, sparsely pubescent. Bracts 1 to 2 at base, upper bract 0.5–3 mm long, 0.5–3 mm wide, broadly ovate, puberulent outside, glabrous inside. Sepals 2, 1–4 mm long, 2–5 mm wide, depressed to broadly ovate, basally fused, pubescent to glabrous outside, glabrous inside, green. Petals 4, 6–15 mm long, 5–8 mm wide, length:width ratio 1.4–1.7, ovate, free, sparsely pubescent to glabrous outside, glabrous inside, light green outside, white inside. Male flowers: stamens ca. 160, 1.1–2 mm long, 0.3–0.5 mm wide, anthers linear, connective prolongation truncate. Female flowers: stamens 1–5, ca. 1 mm long, minute, fertile. Carpels 32 to 100, 2–3.5 mm long, 1–3 mm wide, densely pubescent, free; stigma ca 0.5 mm long, 0.2–1 mm wide. Fruiting pedicel 14–42 mm long, 1–2.5 mm in diameter, glabrous to sparsely pubescent. Monocarps, 2–7, 13.5–37 mm long, 4.5–17 mm wide, length:width ratio 1.1–3, cylindrical, smooth to lumpy, strongly constricted between the seeds, apex rounded to slightly acuminate, sparsely pubescent to glabrous, glossy yellow to red, stipitate; stipe 2.5–14 mm long, 1.5–3 mm wide, sparsely pubescent to glabrous. Seeds 2–6 per monocarp, uniseriate, 13.5–15 mm long, ca. 10 mm wide, ellipsoid, testa smooth, papery, peeling off.

Uvariopsis lovettiana Couvreur & Q.Luke A habit B branch with leaves, upper side C branch with leaves, lower side D leaf base, upper side E leaf base, lower side F flower bud, side view G male flower, top view H female flower, top view. A–F Dagallier 66 G, H Couvreur 97b (type). Photos A–F Léo-Paul Dagallier G, H Thomas Couvreur.
Distribution
Endemic to Somalia-Masai Region: Tanzania.
Habitat and ecology
Submontane mature rain forests. Altitude: 900–1700 m asl.
Phenology
Flowers collected in January, February, May, July, August and from October to December. Fruits collected in February and from June to November.
Notes
Up. lovettiana resemble Up. congensis in having elliptic leaves with acute to decurrent base and attenuate to acuminate apex, and cylindrical monocarps strongly constricted between the seeds. However, Up. lovettiana has generally larger leaves than Up. congensis (15–28 cm long vs. 7–17.7 cm long), longer flower pedicels (14–50 mm long vs. 3–11 mm long), and larger petals (6–15 mm long and 5–8 mm wide vs. 2.5–8 mm long and 1.5–5 mm wide).

Uvariopsis lovettiana Couvreur & Q.Luke A flowering branch B male flower, semi-bottom view C detail of outer petal indumentum D female flower, top view E detail of female flower, three petals removed, side view F carpels G longitudinal section of carpel H male flower, top view I male flower, one petal removed, side view J stamen, lateral view K stamen, back view L stamen, front view M fruit N seed, top view O seed, side view. A–L from Couvreur 97b (type) M–O from Luke 10369. Drawing by Hans de Vries, from “A new species of Uvariopsis (Annonaceae), endemic to the Eastern Arc Mountains of Tanzania” Blumea 55:1, fig. 1, p. 69 (
Preliminary conservation status
This species is endemic from Tanzania. It has previously been assessed preliminarily as Near Threatened based on IUCN criterion B by

Additional specimens examined
Tanzania – Iringa • C. Frimodt-Møller TZ95 (DSM, K), Luhega Forest Reserve, Iringa District, T7; 8°21'S, 35°58'E; alt. 1600 m; 20 Jan. 1997 • C.E. Bracebridge 102 (MO); Kilolo District, Kilombero Nature Reserve, Udekwa Village; 7°48'06'S, 36°30'21'E; alt. 1415 m; 22 Oct. 2009 • D.W. Thomas 3921 (K, MO); Iringa Rural District, Mwanihana Forest Reserve, above Sanje village.Forest on steep slope with small streams and swamps, and patches of elfin forest on ridge top; 7°50'S, 36°55'E; alt. 1400 m; 10 Oct. 1984 • L. Festo 2014 (MO); Kilolo District, Udzungwa Mountains National park, Udzungwa forest, c. 10 km from Msolwa village at source of Msolwa stream; 7°42'11'S, 36°52'23'E; alt. 1350 m; 24 Oct. 2005 • M.A. Mwangoka 8670 (MO); Kilolo District, Uzungwa Scarp Forest Reserve, Idegenda area, E part of Idegenda village; 8°23'S, 35°57'E; alt. 1650 m; 04 Dec. 2012 • W.A. Rodgers 2252 (DSM, K); Iringa Rural District, Udekwa village, forest blowk to East. West of Kilombero F.R; 7°55'S, 36°30'E; alt. 1676 m; Oct. 1982 • W.R.Q. Luke 10369 (K, MO); Kilolo District, Ndundulu Forest reserve, Camp 589 – Camp 590; 7°47'S, 36°29'E; alt. 1440 m; 06 Sep. 2004 • W.R.Q. Luke 6722 (MO); Iringa Rural District, Udzungwa Mountain NP Mt Luhombero Pt 129-131; 7°47'S, 36°32'E; alt. 1440 m; 27 Sep. 2000 • W.R.Q. Luke 7730 (EA, K, NHT), Pt 211; 7°47'S, 36°49'E; alt. 1080 m; 23 Sep. 2001 • W.R.Q. Luke 9195 (EA, K), Pt 370-371; 7°40'S, 36°40'E; alt. 1780 m; 16 Oct. 2002 – Morogoro • A.R. Marshall 1116 (K), Gologolo – PSP10. Uddzungwa Nationa Park. Udzungwa Mountains; 7°41'31.36'S, 36°52'21.02'E; alt. 1772 m; 07 Aug. 2007 • A.R. Marshall 2010 (K); Kilombero District, Ndundulu – Kipunji Plot 1. Kilombero Nature Reserve. udzungwa Mountains; 7°48'17.36'S, 36°30'09.69'E; alt. 1425 m; 30 Jul. 2010 • A.R. Marshall 2114 (K), Udzungwa Forest, Kipunji; 7°48'14.36'S, 36°30'10.01'E; 28 May. 2011 • M.A. Mwangoka 3973 (MO); Mvomero District, Kanga Forest Reserve, Kwamndolwa area; 6°00'S, 37°43'E; 29 Jun. 2005 • M.A. Mwangoka 7178 (MO); Kilosa District, Lumuma Ward, Lunezi Village, Mianzini forest between Lunenzi and Manyomis subvillage; 6°49'18'S, 36°44'36'E; alt. 1530 m; 15 Dec. 2010 • Y.S. Abeid 2625 (MO); Mvomero District, Kanga Forest Resrve, Work Unit 1, North East Transect, 500 m from centre point; 5°59'S, 37°40'E; alt. 900 m; 21 Feb. 2006 – Tanga • A.L. Borhidi 82177 (DSM); Lushoto District, SE corner of Mazumbai University Forest Reserve with ‘Lundgren's plot'; 4°48'S, 38°30'E; alt. 1400 m; 24 Feb. 1982 • B.J. Harris 6257 (DSM), West Usambara Mountains, Mwazumbai; 4°48'S, 38°30'E; alt. 1450 m; 06 Apr. 1972 • C.J. Kayombo 1406 (MO); Korogwe District, T3. Ambangulu Tea Estate, Forest Reserve 5 km W of Ambangulu Tea Factory above Estate road to Makunga; 5°04'48'S, 38°25'05'E; alt. 1300 m; 03 Nov. 1998 • L.-P.M.J. Dagallier 64 (DSM, MPU, WAG); Korogwe District, East Usambaras, Ambangulu, top of the mountain above the tea plantations; 5°04'08.38'S, 38°24'30.45'E; alt. 1316 m; 20 Nov. 2019 • L.-P.M.J. Dagallier 66 (DSM, K, MO, MPU, P, WAG); Korogwe District, East Usambaras, Ambangulu, top of the mountain above the tea plantations; 5°03'57.38'S, 38°23'59.43'E; alt. 1208 m; 20 Nov. 2019.
Uvariopsis noldeae
Type
Angola – Malanje • I. von Nolde 576 (holotype: BM! (BM000554081)), Quela; 9°16'06.17'S, 17°04'12.72'E; alt. 1200 m; Dec. 1938.
Description
Tree height and D.B.H. unknown; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 1–2.5 mm long, ca. 1.5 mm wide, sparsely pubescent to glabrous. Leaf lamina 110–204 mm long, 40–60 mm wide, length:width ratio ca. 3.4, elliptic to obovate, coriaceous, base rounded, apex acuminate, acumen ca. 21 mm long, surface above glabrous, surface below glabrate to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 11–12 pairs, weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers dimorphic, on same individuals (plant monoecious). Flower buds ovoid to conical. Male inflorescences borne on trunk, composed of 1 flower. Peduncle inconspicuous. Flower pedicel 2–4 mm long, ca. 0.5 mm in diameter, pubescent. Bracts 1 to 2 at base, upper bract 0.5–1 mm long, 0.5–1 mm wide, broadly ovate, pubescent outside, glabrous inside. Sepals 2, 1–2 mm long, ca. 2 mm wide, broadly ovate, pubescent outside, glabrous inside. Petals 4, 6.5–10 mm long, 2–3.5 mm wide, length:width ratio 2.8–3.2, ovate, free, pubescent outside, glabrous inside, brownish red outside, cream inside. Stamens numerous (exact number unknown), 0.3–0.6 mm long, 0.1–0.4 mm wide, anthers linear, connective prolongation truncate. Female inflorescences borne on trunk, composed of 1 flower. Flower pedicel 70–83 mm long, ca. 1 mm in diameter, sparsely pubescent. Bracts 1 to 2 at base, upper bract ca. 1.5 mm long, ca. 1.5 mm wide, broadly ovate, pubescent outside, glabrous inside. Sepals 2, 3–4 mm long, ca. 4 mm wide, broadly ovate, pubescent outside, glabrous inside, color unknown. Petals 4, 14–16 mm long, 5–8 mm wide, length:width ratio 1.8–3.2, ovate, free, pubescent outside, glabrous inside, brownish red outside, cream inside. Carpels 40 to 45, ca. 2 mm long, ca. 1 mm wide, velutinous, free, ovules ca. 20 per ovary, biseriate; stigma flat. Fruits unknown.

Distribution
Endemic to Zambezian Region. Known from only one locality in Angola: Quela in Malange region.
Habitat and ecology
Montane rain forests. Altitude ca. 1700 m asl.
Phenology
Flowers collected in December.
Notes
Up. noldeae closely resembles Up. solheidii in having similar leaves and flowers. However, Up. noldeae differs from Up. solheidii in having young branches and petioles sparsely pubescent to glabrous (vs. tomentose to shortly tomentose) and leaf midrib glabrous below (vs. tomentose to glabrous below). Up. noldeae has very dimorphic flowers with pedicels of male flowers 2–4 mm long (vs. 9–30 mm long in Up. solheidii) and pedicels of female flowers 70–83 mm long (vs. 30–198 mm long in Up. solheidii).

Conservation status
This species is known from a single specimen from Angola, collected in 1938, outside any protected area. Angola is one of the least botanically explored countries (
Uvariopsis oligocarpa , sp. nov.
Type
Ghana – Western Region • M.C. Merello 1380 (holotype: MO! (MO3055200); isotype: WAG! (WAG.1570507)), Bia National Forest and Production Reserve. Secondary logging roads west from MIM Timber Company Camp; 6°24'15'N, 3°02'30'W; alt. 140 m; 04 Mar. 1996.
Diagnosis
Up. oligocarpa resembles Up. bisexualis, Up. congensis and Up. zenkeri when sterile, having elliptic leaves generally less than 15 cm long with decurrent base, but it differs clearly when fertile. Up. oligocarpa has unisexual flowers (vs. bisexual flowers in Up. bisexualis) and less carpels (13 to 20 vs. 20 to 40 in Up. congensis), which results in generally 1 to 3 monocarps reaching maturity on the ripe fruit (vs. up to 15 mature monocarps in Up. congensis). The monocarps of Up. oligocarpa are pubescent to sparsely pubescent (vs. glabrate to glabrous in Up. congensis and tomentose in Up. zenkeri), not constricted between the seeds (vs. strongly to slightly constricted between the seeds in Up. congensis and Up. zenkeri), yellow to orange when ripe and brown to dark brown when dry (vs. green to red when ripe and black when dry in Up. congensis), and sessile to very shortly stipitate with stipes shorter than 1 mm (vs. sessile to stipitate with stipes between 0 and 7 mm in Up. congensis). In short, Up. oligocarpa can be distinguished from all the other Uvariopsis species by the combination of the following characters: plant with glabrous branches and elliptic leaves less than 150 mm, unisexual flowers, female flowers with less than 21 carpels, fruits composed of 1–3 monocarps reaching maturity, monocarps pubescent to sparsely pubescent and not constricted between the seeds (Table

Uvariopsis oligocarpa Dagallier & Couvreur A leaves, upper side B leaf, lower side C flower bud, side view D female flower bud, two petals removed, side view E carpels, side view F carpel, top view G receptacle of male flower with stamens, top view H male flower, one petal removed and one petal falling down, side view I fruit with single monocarp, side view, note the hairs on the monocarp J mature fruit, entire (left), longitudinal section (right), side view. A, B, J Koivogui 98 C de Wilde 3656 D, F Merello 1380 H de Koning 2657 I Koivogui 244 E, G van der Laan 640. Photos A, B, I, J Ehoarn Bidault (CC BY-NC-ND 3.0) C–H Léo-Paul Dagallier.
Description
Tree 2–3 m tall, D.B.H unknown; young branches glabrous, old branches glabrous. Petiole 4–6 mm long, 1.7–2.5 mm wide, glabrous. Leaf lamina 90–149 mm long, 32–53 mm wide, length:width ratio 2.2–4, elliptic, papyraceous to coriaceous, base acute to decurrent, apex acuminate, acumen 10–21 mm long, surface above glabrous, surface below glabrous; midrib slightly impressed above, raised below, glabrous above, glabrous below; secondary veins 5–11 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers similar, on same individuals (plant monoecious). Flower buds globose. Male and female inflorescences axillary or sometimes borne on branches, composed of 1 flower. Peduncle inconspicuous. Flower pedicel 2–7 (20) mm long, 1–1.5 mm in diameter, pubescent. Bracts 1 to 3 at base, upper bract 0.5–3 mm long, 0.5–2 mm wide, broadly ovate, pubescent outside, glabrous inside. Sepals 2, 0.7–2.5 mm long, 2–3 mm wide, depressed ovate, free to basally fused, pubescent outside, glabrous inside, color unknown. Petals 4, 4–6.5 (10) mm long, 2.5–6 (10) mm wide, length:width ratio 1–1.4, broadly ovate, free, pubescent outside, glabrous inside, yellowish cream to yellow outside. Male flowers: stamens 300 to 400, 0.5–1.6 mm long, 0.1–0.3 mm wide, anthers linear, connective prolongation truncate. Carpels ca. 20, ca. 1 mm long, ca. 0.5 mm wide, free, stunted, sterile (seen on only one male flower). Female flowers: carpels 13 to 20, 1–2 mm long, 0.5–1.1 mm wide, velutinous, free; stigma ca. 0.1 mm long, ca. 0.5 mm wide, coiled, glabrous. Fruiting pedicel 4–10 mm long, 1–3 mm in diameter, glabrous. Monocarps, 1–3, 25–60 mm long, 11–35 mm wide, length:width ratio 1.7–2.7, cylindrical, smooth, pubescent to sparsely pubescent, green ripening yellow to orange (orange-brown when dry), sessile to very shortly stipitate; stipe 0–1 mm long, 2–3 mm wide, pubescent. Seeds 8–13 per monocarp, biseriate, 8.5–14 mm long, 7–8 mm wide, ellipsoid to semicircular, in an orange pulp.

Distribution
Endemic to Upper Guinean Domain of the Guineo-Congolian Region: Ghana, Ivory Coast, Liberia, Sierra Leone.
Habitat and ecology
Lowland mature or secondary forest. Altitude: 140–285 m asl.
Phenology
Flowers collected from February to June and in November. Fruits collected from January to March and from June to November.
Etymology
The specific epithet refers to the low number of monocarps reaching maturity in this species.
Preliminary conservation status
This species is distributed in West Africa, from Sierra Leone to Ghana. Its EOO is estimated at 147,428 km2 and its AOO at 96 km2. Based on AOO, it would qualify for Endangered EN under B2, the other criteria (a, b, c) are not met. It is thus assigned a preliminary conservation status of Least Concern LC.
Additional specimens examined
Ghana – Brong-Ahafo Region • J.B. Hall 44553 (K), Desiri F.R; 6°45'30.2'N, 2°18'06.66'W; 20 Jun. 1973. Ivory Coast – Abidjan • F.M. van der Laan 640 (U, WAG), Centre Orstom Adiopodonmé, 17 km Dabou rd. from Abidjan. Jardin Botanique; 5°19'30'N, 4°08'W; 29 Jun. 1983 • J.J. Bos 10345 (WAG), Centre Orstom Adiopodonmé, 17 km Dabou road from Abidjan, Jardin Botanique; 5°20'N, 4°07'W; 16 Jun. 1978 – Adzopé • L. Aké Assi 12872 (G), Forêt de la Besso; 6°02'N, 3°41'W; 01 Apr. 1975 – Agboville • A.J.B. Chevalier 16721 (P), Bouroukrou, chemin de fer km 92;6°04'N, 4°18'W; 20 Jan. 1907 • L. Bernardi 8623 (G, K, P), in regione Yapo-Nord, 60–70 km ad septentrionem Abidjan; 5°45'N, 4°07'W; 14 Mar. 1962 – Guiglo • A. Bakayoko 140 (G, P, WAG), Zagné; 6°13'N, 7°29'W; 23 May. 2002 • A.J.B. Chevalier 19308 (P), Keeta, Bassin du moyen Cavally. Pays des Oubi: village de Ke'eta et environs; 5°59'N, 7°28'W; 09 Jul. 1907 • A.J.B. Chevalier 19347 (P), Keeta, Bassin du moyen Cavally. Pays des Oubi: village de Ke'eta et environs; 5°59'N, 7°28'W; 09 Jul. 1907 • L. Aké Assi 12088 (K), Forêt près de Ziriglo; 5°37'N, 7°21'W; 04 Jun. 1973 • L. Aké Assi 13218 (G), entre Taï et Grabo: Troya; 5°42'28.06'N, 7°23'42.49'W; 10 Jan. 1976 • L. Bernardi 8403 (G, K, P), De oppido Tienkulà, ad orientem per 5–9 km in silva alta; 6°08'N, 7°25'W; 02 Mar. 1962 • N. Stäuble 848 (G, MO), Taï; 5°52'N, 7°27'W; Feb. 1982 • N. Stäuble 849 (G, MO), Taï; 5°52'N, 7°27'W; 27 Dec. 1981 • P.R.J. Bamps 2600 (P), Zro; 6°17'45.96'N, 7°34'22.69'W; Apr. 1970 – San-Pédro • C.C.H. Jongkind 4737 (WAG), Forêt Classée Monogaga, just south of Sassandra – San Pedro road; 4°51'48'N, 6°26'30'W; 25 Mar. 2000 – Sassandra • C. Geerling 2328 (WAG), Dakpadou-Sago; 5°16'N, 5°58'W; 29 Mar. 1968 • J. de Koning 2657 (BR, MO, WAG), near Fuyt plantation; 5°03'N, 6°14'W; 11 Nov. 1973 – Soubré • A.J.B. Chevalier 19061 (P), Guideko, Bassin de la moyenne Sassandra entre Guidéko et la Zozro; 5°57'N, 6°35'W; 10 Jun. 1907 • A.J.B. Chevalier 19222 (P), entre le moyen Sassandra et le moyen Cavally; 5°55'59.75'N, 7°03'46.35'W; 06 Jul. 1907 • A.J.B. Chevalier 19253bis (P), entre moy. Sassandra et moy. Cavally; 5°55'59.75'N, 7°03'46.35'W; 06 Jul. 1907 • A.J.B. Chevalier 19282 (P), entre le moyen Sassandra et le moyen Cavally; 5°55'59.75'N, 7°03'46.35'W; 04 Jul. 1907 • M. Scouppe 222 (G), Taï National Park, P.N. de Taï, zone du Centre, secteur Soubré, groupe de transect VH, RL6; 5°43'21.19'N, 7°53'25.45'W; alt. 220 m; 28 Jan. 2010 • P.R.J. Bamps 2507 (P), Guézon – Buyo; 6°30'12.96'N, 7°03'18.22'W; Feb. 1970 – Unknown major area • A. de Rouw 334 (WAG), Pauleoula; 5°49'N, 7°24'W; 08 Dec. 1986. Liberia – Grand Gedeh • J.J.F.E. de Wilde 3656 (K, MA, MO, P, U, WAG), Eastern Province, Putu District. Near Kanweake, a village c. 70 km S. of Chiehn (Zwedru village); 5°30'N, 8°03'W; 26 Mar. 1962 • J.T. Baldwin jr 7091 (K), Zwedru; 6°04'N, 8°08'W; 09 Aug. 1947. Sierra Leone – Eastern Province • B. Saradugu 57 (K, SL, WAG), Gola National Park, central block; 7°39'42.7'N, 10°52'02.6"W; alt. 285 m; 31 Oct. 2013.
Uvariopsis pedunculosa
≡ Tetrastemma pedunculosum Diels, Bot. Jahrb. Syst. 53(3–5): 441 (1915). Type. Cameroon – South Region • G.A. Zenker 3868 (holotype: B! (B 10 0153122); isotypes: BM! (BM000554078), BR! (BR0000008824196, BR0000008824226), E! (E00718574), HBG! (HBG502486), K! (K000199041), M! (M0107937), P! (P00362601, P00362599), US! (US00098850)), Bipinde, Urwaldgebiet; 3°05'N, 10°25'E; 1909.
= Uvariopsis vanderystii Robyns & Ghesq., Ann. Soc. Sci. Bruxelles, Ser. B liii. 64 (1933). Type. Democratic Republic of the Congo – Bandundu • H.J.R. Vanderyst 9973 (holotype: BR! (BR0000008824387)), Kikwit; 5°02'S, 18°49'E; 1921.
Description
Shrub to tree 2–8 m tall, D.B.H 1.5–6.5 cm; young branches pubescent to glabrous, old branches glabrous. Petiole 2–5 mm long, 1.8–3 mm wide, glabrous. Leaf lamina 172–290 mm long, 57–108 mm wide, length:width ratio 2.3–3.9, elliptic to obovate, coriaceous, base acute, apex acuminate, acumen 3–23 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 8–15 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers dimorphic, on same individuals (plant monoecious). Flower buds globose. Male inflorescences borne on trunk, composed of 1 flower. Peduncle inconspicuous. Flower pedicel 14–320 mm long, 0.9–3 mm in diameter, sparsely pubescent to glabrous. Bracts 1 at base and 1 towards the middle or lower half of the pedicel, upper bract ca. 2 mm long, ca. 2 mm wide, broadly ovate, adpressed, semi clasping the pedicel. Sepals 2, 2–7 mm long, 6–7.5 mm wide, very broadly ovate, basally fused, enclosing the petals in bud, sparsely pubescent outside, glabrous inside, dark purplish red. Petals 4, 9–12 mm long, 5–7 mm wide, length:width ratio 1.3–2.2, broadly elliptic to elliptic, fused at base over 20–40 % of their length, pubescent outside, glabrous inside, cream outside, orangish to reddish brown inside. Stamens 650 to 900, 0.5–1 mm long, 0.1–0.3 mm wide, anthers linear, connective prolongation truncate or absent. Female inflorescences borne on trunk, composed of 1 flower. Flower pedicel 80–325 mm long, 1–3 mm in diameter, sparsely pubescent. Bracts 1 at base and 1 towards the middle or lower half of the pedicel, upper bract ca. 2 mm long, ca. 2 mm wide, broadly ovate, adpressed, semi clasping the pedicel. Sepals 2, 6–18 mm long, 9–12 mm wide, very broadly ovate, basally fused, enclosing the petals in bud, sparsely pubescent outside, glabrous inside, dark purplish red. Petals 4, 8–14 mm long, 6–10 mm wide, length:width ratio 1.3–2.2, broadly elliptic to elliptic, fused at base over 20–40 % of their length, pubescent outside, glabrous inside, cream outside, orangish to reddish brown inside. Carpels 50 to 140, 2–4 mm long, 1–1.5 mm wide, pubescent, free; stigma 0.2–0.5 mm long, 0.2–0.5 mm wide, globose. Fruiting pedicel 129–197 mm long, 2–2.2 mm in diameter, pubescent to glabrous. Monocarps, 4–9, 10–17 mm long, 7–11 mm wide, length:width ratio 1.3–1.6 (measures taken from unripe fruits), cylindrical, very verrucose, pubescent, blackish brown, sessile.

Uvariopsis pedunculosa (Diels) Robyns & Ghesq A base of trunk with flower B leaf, lower side C base of leaf, upper side D base of leaf, lower side E male flower, top view F female flower, top view G inflorescence H flower, side view I fruit, top view. A, C, D, F, H Couvreur 594 B, G Bidault 2300 E Couvreur 878 I Couvreur 885. Photos A, C–F, H, I Thomas Couvreur B, G Ehoarn Bidault (CC BY-NC-ND 3.0).
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Cameroon, Democratic Republic of the Congo, Gabon.
Habitat and ecology
Lowland to submontane mature or secondary rain forests. Altitude: 200–1100 m asl.
Phenology
Flowers collected in from February to June and from October to December. Fruits collected in from April to June and in November and December.
Notes
Uvariopsis pedunculosa resembles Up. dioica, Up. guineensis and Up. solheidii in having elliptic to obovate leaves, with acute base and acuminate apex. It has female flower pedicels 80–325 mm long, which is longer than in all Uvariopsis species, except in Up. congolana (200–400 mm long) and Up. solheidii (up to 198 mm long). It differs from Up. congolana in having 4 petals (vs. 3 petals) and verrucose monocarps (vs. longitudinally ridged monocarps). It differs from Up. solheidii in having broadly elliptic to elliptic petals, fused at base over 20–40% of their length (vs. ovate to narrowly ovate and free petals). In the Revision of the Flora of West Tropical Africa, Keay (

Preliminary conservation status
This species has already been assessed (as Uvariopsis vanderystii) as Vulnerable VU under criteria B2ab(iii), mainly because of continuing decline in AOO due to the threat of agricultural expansion (
Additional specimens examined
Cameroon – Littoral • T.L.P. Couvreur 1173 (K, MPU, P, WAG, YA), Mapubi, 30 km before Edea on Yaoundé-Edea road. On forestry road, 5 km direction to Sanaga river; 3°50'44.56'N, 10°23'21.98'E; alt. 219 m; 28 Feb. 2018 – South Region • G.P. Tchouto Mbatchou 3242 (KRIBI, SCA, WAG), Campo-Ma'an area, Massif des Mamelles, along the path to Mamelles highlands; 2°33'57'N, 9°56'58'E; alt. 280 m; 23 Apr. 2001 • G.P. Tchouto Mbatchou ELEX15 (WAG), Campo Ma'an area, Mont d' Elephant, Bidou area towards Hevecam, path to the summit; 2°47'52'N, 10°01'12'E; alt. 180 m; 16 Oct. 2001 • G.P. Tchouto Mbatchou ONOX182 (WAG), Campo Ma'an area, Onoyong, between plots ONO1 and 10; 2°31'39'N, 10°41'49'E; alt. 360 m; 18 Mar. 2001 • G.P. Tchouto Mbatchou ONOX274 (WAG), Campo Ma'an area, Onoyong, between plots ONO1 and 10; 2°31'39'N, 10°41'49'E; alt. 360 m; 18 Mar. 2001 – South-West Region • D.W. Thomas 2756 (K, MO), on the southern slope of Mount Cameroon, above Batoke; 4°05'N, 9°06'E; alt. 500 m; 29 Dec. 1983 • D.W. Thomas 7364 (P), Takamanda Forest Reserve; 6°13'N, 9°26'E; alt. 500 m; 30 Apr. 1987 • G.K. Gottsberger 170307/11 (ULM, WAG), c. 2 km from Banyang Mbo Research Station; 5°08'N, 9°30'E; 17 Mar. 2007 • G.W.J. Mildbraed 10745 (B, K), Likomba – Pflanzung, 15–35 km NE von Victoria; 4°06'N, 9°20'E; alt. 50 m; 03 Dec. 1928 • J.F. Villiers 1427 (P), Massif Ntali, crète sommitale, 30 km SE Mamfé; 5°33'53.69'N, 9°29'26.07'E; alt. 1100 m; 14 Jun. 1982 • R.G. Letouzey 13849 (MO, P), Crète de Nta Ali, entre cotes 1009 et 1202; 30 km SE Mamfe (feuille IGN 1/200.000 Mamfe); 5°33'39.99'N, 9°30'09.81'E; alt. 1266 m; 19 Jun. 1975 • T.L.P. Couvreur 1063 (MPU, WAG, YA), on forest trail, north of Ngomboku village; 4°54'48.49'N, 9°43'28.73'E; alt. 793 m; 06 Apr. 2016 • T.L.P. Couvreur 1066 (MPU, WAG, YA), on forest trail, north of Ngomboku village; 4°54'38.74'N, 9°43'48.18'E; alt. 815 m; 06 Apr. 2016 • T.L.P. Couvreur 517 (MPU, YA); Fako, on trail trough palm oil plantation, 3 km before lava flow and Seme Beach hotel when coming from Limbe; 4°04'28.6'N, 9°05'06.64'E; alt. 479 m; 18 Oct. 2013 – Unknown major area • W.G. Gosline 244 (K, MO, P, WAG, YA), Kupe-Muanenguba Division, Ajang saprophyte plot; 4°46'N, 9°41'E; alt. 950 m; 01 Dec. 1999. Equatorial Guinea – Centro Sur • B. Senterre 2989 (BRLU), SO du Parc National de Monte Alén, 2 km au NE du site de traversée du rio Uolo pour aller aux cataractas; 1°36'33.62'N, 10°05'32.96'E; alt. 750 m; 23 Jun. 2002. Gabon – Estuaire • T.L.P. Couvreur 602 (MPU, YA), Crystal Mountains, on trail behind the dam at Tchimbélé; 0°37'02.43'N, 10°24'41.26'E; alt. 617 m; 16 Nov. 2013 • T.L.P. Couvreur 607 (MPU, YA), Crystal Mountains, on trail befind tunnel leading under dam; 0°36'47.64'N, 10°24'10.96'E; alt. 492 m; 16 Nov. 2013 – Nyanga • J.L.C.H. van Valkenburg 2940 (BR, LBV, WAG), Moukalaba Doudou, national park, south of the road to Kachimba; 2°41'36'S, 10°27'06'E; alt. 500 m; 22 Feb. 2004 – Ogooué-Ivindo • E. Bidault 2300 (BR, BRLU, LBV, MO, MO, P, WAG), concession forestière Rougier-Ivindo; 0°06'23.9'N, 12°21'06.3'E; alt. 380 m; 29 Oct. 2015 • M.S.M. Sosef 2241 (BR, K, LBV, MO, WAG), c. 30 km down the Ivindo River from the IRET Research Station, SW of Makokou; 0°21'30'N, 12°38'42'E; alt. 350 m; 07 Nov. 2005 – Ogooué-Lolo • G.M.P.C. Le Testu 8525 (P), region de Lastoursville, Lastoursville; 0°50'N, 12°42'E; 18 Nov. 1930 – Woleu-Ntem • J.M. Reitsma 2554 (LBV, MO, WAG), Inventory Oveng; primary rain forest, ca 25 km WSW of Mintsic; 0°44'N, 11°22'E; 08 Nov. 1986 • J.M. Reitsma 891 (LBV, MO, WAG), chantier Rougier-Océan, Oveng; 0°40'N, 11°22'E; alt. 760 m; 08 May. 1985 • T.L.P. Couvreur 594 (MPU, YA), 40 km from Sam, on raod main road to Medouneu; 0°42'14.82'N, 10°20'53.39'E; alt. 589 m; 14 Nov. 2013 • T.L.P. Couvreur 878 (LBV, WAG, YA), on road from Mitzic to Lalara (N2), just after the bridge over the Lara, c.500 m in forest; 0°36'15.66'N, 11°29'12'E; alt. 574 m; 15 Nov. 2015 • T.L.P. Couvreur 885 (LBV, WAG, YA), c.15 km south of Mitzic (N2), in Foreex logging conssession, 5 km after the bridge after the main base camp (Saint Germain, c.25 km from road N2), on abandoned logging road left to the main road; 0°41'51.34'N, 11°40'50.29'E; alt. 531 m; 16 Nov. 2015.
Uvariopsis solheidii
≡ Tetrastemma solheidii De Wild., Ann. Mus. Congo Belge, Bot. sér. 5, 3(1): 85 (1909). Type. Democratic Republic of the Congo – Orientale • A.F. Solheid 96 (holotype: BR! (BR0000008824240)); Banalia, Env. de Yambuya; 1°16'N, 24°33'E; 1906.
= Uvariopsis batesii Robyns & Ghesq., Ann. Soc. Sci. Bruxelles, Ser. B liii. 320 (1933). Type. Cameroon – South Region • G.L. Bates 1367 (holotype: BM! (BM000554077)), Bitye; 3°00'41.45“N, 12°21'04.18"E; 1919.
= Uvariopsis letestui Pellegr.; syn. nov. concerning Uvariopsis letestui var letestui (see details under this variety).
Description
Shrub to tree 1.5–8 m tall, D.B.H 2–6 cm; young branches tomentose to shortly tomentose, old branches glabrous. Petiole 2–5 mm long, 1.5–3 mm wide, tomentose to glabrous. Leaf lamina 131–260 (285) mm long, 40–93 mm wide, length:width ratio 2.1–3.9, elliptic to obovate, coriaceous, base acute to rounded to subcordate, apex attenuate to narrowly acuminate, acumen 4–20 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, tomentose to glabrous below; secondary veins 7–13 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers dimorphic, on same individuals (plant monoecious, but individuals with only pistillate flowers were also reported; e.g. Le Testu 8458). Flower buds long ovoid to conical. Male inflorescences borne on small thickenings of the trunk, mainly between the base towards 50 cm, up to ca. 2.50 m, above the female inflorescence, composed of 1 to 2 flowers. Peduncle inconspicuous. Flower pedicel 9–30 mm long, 0.5–1 mm in diameter, pubescent to sparsely pubescent. Bracts 1 to 2 at base and none to 1 towards the lower half of the pedicel, upper bract 1–2 mm long, 1.5–3 mm wide, broadly ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 1.5–2 mm long, 1.5–2 mm wide, broadly ovate, free, pubescent outside, glabrous inside, brown to red. Petals 4, 5–10 mm long, 2.5–3.5 mm wide, length:width ratio 2–3.2, ovate to narrowly ovate, free, straight to curving outward at anthesis, pubescent to sparsely pubescent outside, glabrous inside, brown to light red outside, dull pink to dark purplish red inside. Stamens ca. 300, 0.2–0.5 mm long, 0.2–0.3 mm wide, anthers linear, connective prolongation truncate. Female inflorescences borne on small thickenings of the trunk, mainly between the base towards ca. 50 cm, up to ca. 2.50 m, below the male inflorescence, composed of 1 to 2 flowers. Flower pedicel 30–198 mm long, 1–1.5 mm in diameter, pubescent to glabrous. Bracts 1 to 2 at base and none to 1 towards the lower 30% of the pedicel, upper bract 1–2 mm long, 1.5–3 mm wide, broadly ovate, adpressed, semi clasping the pedicel, pubescent outside, glabrous inside. Sepals 2, 1.5–4 mm long, 2–5 mm wide, broadly ovate, free, pubescent outside, glabrous inside, brown to red. Petals 4, 7.5–17 mm long, 3.5–7 mm wide, length:width ratio 1.3–3.6, ovate to narrowly ovate, free, straight to curving outward at anthesis, pubescent outside, glabrous inside, brown to light red outside, dull pink to dark purplish red inside. Carpels 30 to 65, 1–3.5 mm long, 1–1.5 mm wide, velutinous, free; stigma globose. Fruiting pedicel 30–176 mm long, 1–2 mm in diameter, pubescent to glabrous. Monocarps, 1–5, 21–70 mm long, 12–30 mm wide, length:width ratio 1.8–3 (measures taken from both dried fruits and their associated specimen label, note that monocarps seem to sink when drying), cylindrical, with 4–6 longitudinal ridges and several transversal ridges or wrinkles, sparsely pubescent to glabrous, bright red, subsessile; stipe 0–4 mm long, 4–5 mm wide, pubescent to glabrous. Seeds 2–10 per monocarp, uniseriate to biseriate, 11–18 mm long, 10–12 mm wide, ellipsoid.

Uvariopsis solheidii (De Wild.) Robyns & Ghesq A leaf, upper side B leaf base, upper side C leaf base, lower side D flower buds, side view, var. solheidii (top), var. letestui (bottom) E inflorescence, side view F male flowers, top view G drawing of: flowers with details (left), young fruits (top right), young monocarp (bottom right). A–C, D (bottom) Couveur 550 D (top), E, F Couvreur 855 G Hallé 3474. Photos Thomas Couvreur. Drawing Nicolas Hallé, part of specimen Hallé 3474 (P).
Distribution
Element of the Lower Guinean Domain and Congolia Domain of the Guineo-Congolian Region: Cameroon, Central African Republic, Democratic Republic of the Congo, Gabon, Republic of the Congo.
Habitat and ecology
Lowland to submontane mature or old secondary rain forests. Altitude: 0–1200 m asl.
Phenology
Flowers collected from January to May, in November and December. Fruits collected in February, from April to August, in October and December.
Vernacular names
Central African Republic: ‘Molo-Mobay’ in Bissongo (Tisserant 2422), ‘Ita ti ngaingai’ in Bambindjere (Fay 8384). Gabon: ‘Héli’ in Bakota (Hallé 3029).
Notes
Up. solheidii resembles Up. dioica, Up. guineensis, and Up. pedunculosa in having elliptic to obovate leaves, with acute to rounded base and attenuate to acuminate apex. Leaf base of Up. solheidii can also be subcordate, and differs from these species in having young branches and petioles tomentose to shortly tomentose (vs. pubescent to glabrous) and in having cylindrical monocarps with longitudinal and transversal ridges (vs. smooth in Up. dioica, with a single longitudinal ridge in Up. guineensis and verrucose in Up. pedunculosa).

Uvariopsis solheidii var. solheidii A male inflorescence B female flower, one petal removed, side view C petal of female flower, inner view D carpel, side and front views, detail of ovules E male flower, one petal removed, side view F petal of male flower, inner view G stamen, front and side views – var. letestui (Pellegr.) Dagallier & Couvreur H flower bud I petal of female flower, inner view J female flower, all four petals removed, side view K carpel, front view and detail of ovules L male flower, one petal removed, side view M stamen, front view N fruit O longitudinal section of A monocarp Uvariopsis pedunculosa (Diels) Robyns & Ghesq P female inflorescence Q female flower, one petal removed, side view R petal of female flower, inner view S carpel, side view and detail of ovules. A from Tisserant 2422 B–G from Tisserant 804 H–M from Hallé 3060 N, O from Hallé 2975 P–S from Le Testu 8525. Drawings by Hélène Lamourdedieu, from
Here we make the two names Uvariopsis solheidii (De Wild.) Robyns & Ghesq. and Uvariopsis letestui Pellegr. synonyms. They have usually been distinguished based on the shape of their petals (narrowly ovate petals in Ud. solheidii vs. elliptic-ovate petals in Ud. letestui), the position of the bracts along the flowering pedicel, and the length of the flower pedicels (longer in Ud. solheidii than in Ud. letestui, but with an overlap) (
Conservation status
This species is widespread in central Africa. It has been assessed as Least Concern LC (Botanic Gardens Conservation International and IUCN SSC Global Tree Specialist Group 2019b). Here its EOO is estimated at 1,429,739 km2 and its AOO at 132 km2.
Uvariopsis solheidii var. letestui , stat. nov.
≡ Uvariopsis letestui Pellegr. syn. nov., Bull. Soc. Bot. France 95: 139 (1948). Type. Gabon – Ogooué-Lolo • G.M.P.C. Le Testu 8458 (holotype: P! (P00362609), sheet here designated; isotypes: BM! (BM000554079, BM000554080), P! (P00362607)), region de Lastoursville, Koulamotou; 1°07'S, 12°30'E; 20 Oct. 1930.
Description
Male flowers with petals 5–7 mm long, ca. 3 mm wide, length:width ratio ca. 2, ovate, free, straight at anthesis. Female flowers with pedicel 30–110 mm long; petals 7.5–10 mm long, 4.5–7 mm wide, length:width ratio 1.3–1.8, ovate, free, straight at anthesis. Fruiting pedicel 30–110 mm long.
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Gabon.
Habitat and ecology
Lowland to submontane mature rain forests. Altitude: 280–950 m asl.
Notes
Up. solheidii var. letestui differs from the type variety in having slightly different flowers. The male flowers of Up. solheidii var. letestui have ovate petals 5–7 mm long, with a length:width ratio of ca. 2 (vs. ovate to narrowly ovate petals, 7–10 mm long with a length:width ratio of ca. 3). The female flowers of Up. solheidii var. letestui have generally smaller flower pedicels (30–110 mm long vs. 63–198 mm long), and ovate petals 7.5–10 mm long, with a length:width ratio between 1.3 and 1.8 (vs. ovate to narrowly ovate petals, 9–17 mm long with a length:width ratio between 1.8 and 3.6). Both male and female flowers of Up. solheidii var. letestui have straight petals at anthesis (vs. curved outward at anthesis in the type variety).
Additional specimens examined
Gabon – Ngounié • T.O.B.E.B. Stévart 3993 (BR, LBV, MO, WAG), Birougou National Park, Massif du Chaillu, 40 km SE of Mbigou, 5.5 km E of Moukimbi village. MBG transect T59; 2°02'39'S, 12°14'01'E; alt. 780 m; 16 Feb. 2011 – Ogooué-Ivindo • J. Florence 1578 (P), Station d'Ipassa, 10 Km S de Makokou; 0°30'N, 12°47'E; alt. 500 m; 22 Jan. 1979 • N. Hallé 2975 (K, P, P), Bélinga; 1°05'N, 13°08'E; alt. 950 m; 03 Nov. 1964 • N. Hallé 3006 (P), Bélinga; 1°05'N, 13°08'E; 04 Nov. 1964 • N. Hallé 3029 (P), Bélinga; 1°05'N, 13°08'E; 05 Nov. 1964 • N. Hallé 3060 (P), Bélinga; 1°05'N, 13°08'E; 06 Nov. 1964 – Ogooué-Maritime • T.L.P. Couvreur 550 (MPU, YA), along road between Mandji and Rabi. About km 19 from Mandji; 1°46'05.10'S, 10°17'01.5'E; alt. 282 m; 08 Nov. 2013.

Uvariopsis solheidii var. solheidii
Description
Male flowers with petals 7–10 mm long, 2.5–3.5 mm wide, length:width ratio ca. 3, narrowly ovate, free, curving outward at anthesis. Female flowers with pedicel 63–198 mm long; petals 9–17 mm long, length:width ratio 1.8–3.6, narrowly ovate, free, curving outward at anthesis. Fruiting pedicel 53–176 mm long.
Distribution
Element of the Lower Guinean Domain and Congolia Domain of the Guineo-Congolian Region: Cameroon, Central African Republic, Democratic Republic of the Congo, Gabon, Republic of the Congo.
Habitat and ecology
Lowland to submontane mature or old secondary rain forests. Altitude: 0–1200 m asl.
Additional specimens examined
Cameroon – Central Region • B.-A. Nkongmeneck 273 (P), Mt Ngoro à 58 km de Lint. Feuille IGN 1/200.000 LINTE; 5°04'53.52'N, 11°16'56.76'E; 17 Apr. 1982 • J.-M. Onana 2848 (K, SCA, WAG, YA), Mefou Proposed National Park, Ndanan 1, River N Didoumou; 3°37'N, 11°34'E; alt. 720 m; 26 Mar. 2004 • M.R. Cheek 11606 (K, SCA, WAG, YA), Mefou National Park Ndanan 1. Trail to Oncoba flag. (l after bridge); 3°37'13'N, 11°35'04'E; alt. 710 m; 10 Mar. 2004 – East Region • J.N. Asonganyi 310 (P), between Somalomo and Milon, 69 km SE Akonolinga, Map IGN 1/200.000 Akonolinga; 3°21'N, 12°44'E; 18 Jun. 1981 – South Region • A. Koufani 154 (P), Ngongonjie hill, near Akonetye; 2°40'N, 12°52'E; alt. 650 m; 30 Aug. 1978 • G.L. Bates s.n (BM), Bitye, Yaunde, Cameroons; 3°00'41.45'N, 12°21'04.18'E; 1919 • G.P. Tchouto Mbatchou 2843 (KRIBI, WAG), Efoulan, Ongongo and Nkolomekok hills in Akom II area; 2°44'43'N, 10°31'48'E; alt. 1000 m; 25 Apr. 2000 • G.P. Tchouto Mbatchou 3089 (KRIBI, SCA, WAG, YA), Efoulan; 2°44'45'N, 10°32'52'E; alt. 960 m; 04 Dec. 2000 • G.P. Tchouto Mbatchou EGONX191 (WAG), Campo Ma'an area, Akom II, Efoulan and Egongo hills; 2°44'46'N, 10°32'53'E; alt. 840 m; 06 Dec. 2000 • J.F. Villiers 893 (P), colline de Nkoltsia, 23 km NW de Bipindi; 3°10'30'N, 10°16'30'E; 27 Apr. 1974 • J.J.F.E. de Wilde 8373B (WAG), East face of Zingui hill, 21 km on the road from Ebolowa to Kribi (counted from the crossing at Ebolowa); 2°51'N, 10°59'E; alt. 760 m; 21 Jul. 1975. Central African Republic – Lobaye – Équipe C. Tisserant 2422 (BM, P, WAG); Mbaïki, Station de Boukoko; 3°54'N, 17°56'E; 26 Apr. 1952 • Équipe C. Tisserant 804 (K, P, WAG); Mbaïki, Forèt Soms bois; 3°54'N, 17°56'E; 23 Mar. 1948 – Sangha-Mbaéré • J.M. Fay 8384 (K, MO, P, WAG), Ndakan Gorilla Study Area; 2°20'N, 16°11'E; alt. 380 m; 26 May. 1988. Democratic Republic of the Congo – Bandundu • J.L.P. Louis 15474 (BM, K, P); 4°55'54'S, 18°44'42'E; alt. 470 m; 05 Dec. 1939 – Maniema • R.E. Gereau 7613 (MO), Secteur Bangengele; parc National proposé de la Lomani, ca. 330 m à l'Est du camp T7 Lomami et 1.3 km à l'Est de la rivière Lomami sur la piste au camp Kaka Barkata; 2°41'42'S, 25°05'18'E; alt. 450 m; 28 Apr. 2015 – Orientale • J. Bokdam 3674 (KIS, WAG); Banalia, 22 km along road from Kisangani to Bengamisa; 0°43'N, 25°12'E; 16 Nov. 1972 • J. Bokdam 4153B (WAG); Banalia, 23 km along road from Kisangani to Bengamisa; 0°46'N, 25°13'E; 31 May. 1973 • J.-B. Ndjango 549 (EPU, K, K, YBI); Haut-Uele, Asonga Hill, c. 75 km South of Isiro. Outskirts of the Ituri forest. Temporary camp; 2°06'35'N, 27°31'17'E; alt. 960 m; 24 Jul. 2011 • J.L.P. Louis 13439 (K); Isangi, Bassao (île), Un peu en aval de Loléko; 0°54'N, 24°12'E; 27 Jan. 1939 • J.L.P. Louis 14540 (BM, K, MO, P); Isangi, Yangambi; 0°46'N, 24°27'E; alt. 470 m; 09 Sep. 1939 • J.L.P. Louis 8357 (BR, MO, P); Isangi, Yangambi; 0°46'N, 24°27'E; alt. 470 m; 08 Mar. 1938. Gabon – Estuaire • M.E. Leal 268 (LBV, WAG), Monts de Cristal, Mbe National Parc, south of Mont Mbilan; 0°27'40'N, 10°15'30'E; alt. 550 m; 12 Feb. 2005 – Ngounié • M.E. Leal 1906 (LBV, MO, WAG), Bouvala hills. Col between the two summits; 1°37'15'S, 11°45'45'E; alt. 995 m; 06 Oct. 2007 – Ogooué-Ivindo • A.H. Gentry 33052 (MO, P), Makokou. Transect 1; 0°34'N, 12°52'E; alt. 480 m; 01 Jul. 1981 • J. Florence 1158 (P), Station d'Ipassa, 10 km S de Makokou; 0°30'N, 12°47'E; alt. 500 m; 04 May. 1978 • N. Hallé 3474 (K, P, P, U), Bélinga. Route de Mvadi; 1°05'N, 13°08'E; alt. 800 m; 12 Dec. 1964 – Woleu-Ntem • MINKébé Series R67 (WAG), Minkébé area, 10 x 10 m inventory plot R, placed 2–12 m south at 1380–1390 m on transect A; 1°30'N, 12°48'E; Apr. 1990 • MINKébé Series W289 (WAG), crest to the south of Mount Minkébé; 1°35'N, 12°52'E; 10 May. 1990 • T.L.P. Couvreur 856 (LBV, WAG, YA), Oyem, on road to Mbolonzok (off main road to Mongono and Equatorial Guinea); 1°29'21.34'N, 11°27'58.98'E; alt. 675 m; 13 Nov. 2015 – Unknown major area • N. Hallé 2975 (U). Republic of the Congo – Sangha • S.T. Ndolo Ebika 294 (E, IEC, WAG), Nouablé-Ndoki National Park, Goualougo Study Site, 37.53 km E-SE de Bomassa; 2°11'21.34'N, 16°31'24.35'E; alt. 383 m; 18 Jan. 2008 • S.T. Ndolo Ebika 659 (E, IEC, WAG), Périphérie du Nouable Ndoki National Park: 19 Km E de Bomassa; 2°12'23.29'N, 16°20'19.93'E; alt. 381 m; 06 Jun. 2011.
Uvariopsis submontana
Type
Cameroon – South-West Region • D. Kenfack 1334 (holotype: YA; isotypes: K! (K000683145), MO, SCA), Rumpi Hills; 4°57'N, 9°02'E; alt. 800 m; 06 Feb. 2000.
Description
Tree 4.5–25 m tall, D.B.H 5–30 cm; young branches sparsely pubescent to glabrous, old branches glabrous. Petiole 3–8 mm long, 2.5–4 mm wide, sparsely pubescent to glabrous. Leaf lamina 160–380 mm long, 50–110 mm wide, length:width ratio 2.7–4.3, obovate, coriaceous, base rounded to subcordate, apex attenuate to acuminate, acumen 8–30 mm long, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins 9–25 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers slightly dimorphic, on same individuals (plant monoecious). Flower buds ovoid to conical. Male inflorescences borne in dense clumps on thickenings at base of the trunk, sparser above 2 m., above the clumps of female inflorescences, mainly between the base and the lower 5 m of the trunk, composed of 6 to 50 flowers. Peduncle inconspicuous. Flower pedicel 25–50 mm long, ca. 1 mm in diameter, pubescent to sparsely pubescent. Bracts 2 to 4 at base, upper bract 1–1.6 mm long, 1–1.6 mm wide, broadly ovate, pubescent outside, glabrous inside. Sepals 2, 5–11 mm long, 6–12 mm wide, broadly ovate, pubescent to sparsely pubescent outside, glabrous inside, dark red. Petals 4, 8–19 mm long, 4–8 mm wide, length:width ratio 1.8–4.7, ovate to narrowly ovate, free to fused at base over 30% of their length, pubescent outside, glabrous inside, pinkish to dark red outside. Stamens 100 to 200, 0.4–0.7 mm long, 0.3–0.4 mm wide, anthers linear, connective prolongation truncate. Female inflorescences borne in dense clumps on thickenings at base of the trunk, below the clumps of male inflorescences, composed of 6 to 50 flowers. Flower pedicel 24–60 mm long, ca. 1 mm in diameter, pubescent to sparsely pubescent. Bracts 2 to 4 at base, upper bract 1–1.6 mm long, 1–1.6 mm wide, broadly ovate, pubescent outside, glabrous inside. Sepals 2, 6–8 mm long, 6–9 mm wide, broadly ovate, pubescent to sparsely pubescent outside, glabrous inside, dark red. Petals 4, 15–17 mm long, 5–9 mm wide, length:width ratio 1.8–4.7, ovate to narrowly ovate, free to fused over 30% of their length, pubescent outside, glabrous inside, pinkish to dark red outside. Carpels 50 to 100, 1.5–3.5 mm long, 0.8–1.5 mm wide, pubescent, free; stigma ca. 0.5 mm long, ca. 0.2 mm wide, globular, glabrous. Fruiting pedicel 25–90 mm long, 3–7 mm in diameter, glabrous. Monocarps, 3–25, 17–80 mm long, 13–55 mm wide, length:width ratio, ovoid to cylindrical, constricted between seeds in dried specimens, sparsely pubescent to glabrous, pale green ripening dark yellow, sessile. Seeds 6–12 per monocarp, biseriate, 18–25 mm long, 8–13 mm wide, ellipsoid.

Uvariopsis submontana Kenfack, Gosline & Gereau A trunk with clumps of inflorescences B leaf, upper side C leaf base lower side D leaf base, upper side E female flower, two petals removed, side view F flower, top view G clumps of inflorescences on the trunk H male flower, two petals removed, side view I male flower, top view. A–I Couvreur 627. Photos Thomas Couvreur.
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region. Only known from the South-West and Littoral Regions in Cameroon, mainly in Mont Koupé and Ebo National Park.

Uvariopsis submontana Kenfack, Gosline & Gereau A leaves B female flower bud, side view C male flower, side view D male flower, one petal removed, side view E inner surface of sepal F outer surface of sepal G female flower, all four petals removed, side view H carpel, lateral view I infructescence J longitudinal section of monocarp K transversal section of monocarp. A–H from Kenfack 1334 (type) I–K from Kenfack 1373. Drawings by David Kenfack, from “The Genus Uvariopsis (Annonaceae) in Tropical Africa, with a Recombination and One New Species from Cameroon”, Novon 13:4, Figure
Habitat and ecology
Submontane mature or old secondary rain forests. Soil: rocky, humic combisol. Altitude: 900–1300 m asl.
Phenology
Flowers collected from January to May and in December.
Vernacular names
Cameroon: ‘Michile’ in Bakossi (Cheek 7131).
Notes
This species resembles Up. bakeriana, Up. citrata and Up. korupensis in having large obovate leaves (up to 38 cm long), with rounded to cordate bases. It differs from Up. bakeriana and Up. citrata in having longer flower pedicel (24–60 mm vs. 0–8 mm). It is very similar to Up. korupensis, but has generally smaller leaves (16–38 cm vs. 28–62 cm in Up. korupensis). The female flowers of Up. submontana have longer sepals (6–8 mm long vs. 3–5 mm long) and smaller petals (15–17 mm long vs. 15–35 mm long) than Up. korupensis, which results in petals less than 3 times longer than the sepals (vs. petals more than 3 times longer than the sepals). Petals of Up. submontana are pinkish to dark red inside and outside whereas petals of Up. korupensis are cream to brownish outside and cream to pinkish inside (Table

Conservation status
This species is endemic to Cameroon and has been evaluated as Endangered EN B1ab(iii)+2ab(iii) and mainly threatened by the expansion of agriculture (
Additional specimens examined
Cameroon – Littoral • D. Kenfack 1602 (MO), Nkam, Yingui, Bataba. PE80; 4°32'N, 10°18'E; alt. 780 m; 20 Feb. 2002 • T.L.P. Couvreur 627 (MPU, YA), Ebo Wildlife Reserve, Djuma permanent camp. On east trail; 4°21'39.35'N, 10°15'07.7'E; alt. 957 m; 15 Feb. 2013 – South-West Region • J.F. Villiers 2490 (P), 6 km WNW Bomana, 34 km NW Limbé; 4°30'24.99'N, 9°00'50.26'E; 15 Dec. 1984 • M.R. Cheek 7034 (K, WAG, YA), Mount Kupe. Kupe village. Esense river near farm of Philip Taza; 4°46'N, 9°41'E; alt. 1000 m; 19 Jan. 1995 • M.R. Cheek 7131 (K, WAG), Kupe village. Hunters path from Kupe village to the mountain top, running to S and parallel with Esense river (now dry). Towards village largely coffee farms with shade trees. Grid ref. from Magellan reading where path enters forest; 4°47'N, 9°41'E; alt. 840 m; 24 Jan. 1995 • P. Lane 490 (K, SCA, WAG, YA), Mount Kupe. Ndum. Forest trail 2 km S from Etube-Tape village; 4°51'N, 9°42'E; alt. 1100 m; 01 Feb. 1995 • S. Cable 1221 (K, MO, SCA, WAG, YA), Mt Kupe, Nyasoso, Shrike trail; 4°49'00.12'N, 9°42'E; alt. 1100 m; 08 Feb. 1995 • S. Cable 2736 (K, MO, P, SCA, WAG, YA), Kupe-Muanenguba Division. Path on to ridge between Daniel's saprophyte site and Kupe rock; 4°47'N, 9°44'E; alt. 1000 m; 30 May. 1996 • T.L.P. Couvreur 1059 (MPU, WAG, YA), Nyasoso village, on max's trail to Mt Kupe; 4°49'47.02'N, 9°41'34.79'E; alt. 1024 m; 05 Apr. 2016 • T.L.P. Couvreur 965 (MPU, WAG, YA), Rumpi mountains, forest trail, ca. 5 km after Dikome Balue village, ca. 40 km north of Kumba; 4°56'10.37'N, 9°14'30.34"E; alt. 1418 m; 10 Jan. 2016.
Uvariopsis zenkeri
Type
Cameroon – South Region • G.A. Zenker 1117 (holotype: B! (B 10 0153124); isotypes: BM! (BM000554075), G! (G00420324), GOET! (GOET005734), HBG! (HBG502515), K! (K000199042), LE! (LE00012462), M! (M0107936), P! (P00362604), S! (S07-11029), WU! (WU0025790)), Bipinde, Urwaldgebiet; 3°05'N, 10°25'E; 1896.
Description
Shrub to tree 2–5 m tall, D.B.H 1.5–2 cm; young branches densely pubescent to pubescent, old branches glabrous. Petiole 2–5.5 mm long, 1–1.5 mm wide, pubescent to glabrous. Leaf lamina 110–158 mm long, 36–58 mm wide, length:width ratio 2.1–3.4, elliptic to obovate, papyraceous to coriaceous, base acute to decurrent, apex acuminate, acumen 10–24 mm long, surface above glabrous, surface below puberulent to glabrous when young, glabrous when old; midrib impressed above, raised below, glabrous above, pubescent to glabrous below; secondary veins 6–13 pairs, brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unisexual, male and female flowers similar, on same individuals (plant monoecious). Flower buds globose to subovoid. Male and female inflorescences borne on trunk, old branches or old axillary, composed of 1, (sub)sessile flower. Peduncle inconspicuous. Flower pedicel 0–7 mm long, 1–1.5 mm in diameter, pubescent. Bracts 1 to 2 (5 in bud) at base, upper bract 1–4 mm long, 2–5 mm wide, ovate, pubescent outside, glabrous inside. Sepals 2, 3–7 mm long, 4.5–7 mm wide, broadly ovate, basally fused, pubescent outside, glabrous inside, brown. Petals 4, 6–13 mm long, 4–7 mm wide, length:width ratio 1.5–2.2, ovate to triangular, fused at base over ca. 50 % of their length, pubescent outside, glabrous inside, brown outside. Male flowers: stamens 100 to 150, ca. 0.5 mm long, ca. 0.3 mm wide, anthers linear, connective prolongation truncate or absent. Female flowers: carpels 13 to 22, 1.5–3 mm long, 1–1.5 mm wide, velutinous, free; stigma 0.1–0.5 mm long, ca. 1 mm wide, coiled, glabrous. Fruiting pedicel 2–10 mm long, ca. 2.5 mm in diameter, pubescent. Monocarps, 1–3, ca. 37 mm long, ca. 17 mm wide, length:width ratio ca. 2.2, cylindrical, slightly veined and constricted between the seed when dried, tomentose, brown, subsessile; stipe ca. 1 mm long, ca. 2 mm wide, tomentose. Seeds 10–12 per monocarp, biseriate, ca. 13 mm long, ca. 7 mm wide, ellipsoid.

Uvariopsis zenkeri Engl A leaf, upper side B male flowers on brnch C male flower, one petal removed D stamen E female flower, one petal removed F carpel, front and side view, detail of ovules. Uvariopsis congensis Robyns & Ghesq. G flowering branch with male flower bud H male flower, one petal removed I stamen J flowering branch with female flower K female flower, one petal removed L carpel, side view and detail of ovules M fruit A–E from Zenker 1117 F from Zenker 63 G–L from Tisserant 1363 M from Letouzey 5494. Drawings by Hélène Lamourdedieu, Publications Scientifiques du Muséum national d’Histoire naturelle, Paris.
Distribution
Endemic to Lower Guinean Domain of the Guineo-Congolian Region: Cameroon.
Habitat and ecology
Lowland mature or secondary rain forests. Soil: rocky, humic, volcanic. Altitude: 0–m asl.
Phenology
Flowers and fruits collected in April.
Notes
This species resembles Up. bisexualis, Up. congensis and Up. oligocarpa in having elliptic leaves generally less than 16 cm long with decurrent base. It differs from these species in having densely pubescent to pubescent young branches (vs. pubescent to glabrous). It also differs from Up. bisexualis with its unisexual flowers (vs. bisexual flowers). Up. zenkeri has less carpels (13 to 22 vs. 20 to 40 in Up. congensis), which results in generally 1 to 3 monocarps reaching maturity on the ripe fruit (vs. up to 15 mature monocarps in Up. congensis). The monocarps of Up. zenkeri are tomentose (vs. glabrate to glabrous in Up. congensis and pubescent to sparsely pubescent in Up. oligocarpa), slightly constricted between the seeds (vs. not constricted between the seeds in Up. oligocarpa) (Table

Conservation status
This species is endemic to Cameroon. It occurs in nine locations, only four of which are situated in protected areas (Campo-Ma’an National Park, Ebo National Park, Mont Cameroun National Park, Takamanda National Park). It has been previously assessed as Vulnerable VU under criteria B2ab(i,ii,iii,iv,v) (
Additional specimens examined
Cameroon – South Region • G.A. Zenker 3045 (B, BM, G, K, L, M, MA, WAG), Bipindi; 3°05'N, 10°25'E; 1904 • G.A. Zenker 3228 (G, L, M), Bipinde, Urwaldgebiet; 3°05'N, 10°25'E; 1904 • G.A. Zenker 515 (B, G, G, M, MA, MO, WAG), Bipindi; 3°05'N, 10°25'E; Mar. 1914 • G.P. Tchouto Mbatchou 2846 (KRIBI, WAG), Efoulan, Ongongo and Nkolomekok hills in Akom II area; 2°44'43'N, 10°31'48'E; alt. 1000 m; 25 Apr. 2000 • G.P. Tchouto Mbatchou 3252 (KRIBI, SCA, WAG), Campo-Ma'an area, Massif des Mamelles, Mamelles highlands; 2°33'57'N, 9°56'58'E; alt. 280 m; 23 Apr. 2001 • T.L.P. Couvreur 707 (MPU, WAG, YA), Campo Ma'an National Park, 11 km on trail from Ebinanemeyong village, on road, 7 km from Nyabessan to Campo town; 2°28'24.59'N, 10°20'41.79'E; 14 Feb. 2015 – South-West Region • D.W. Thomas 3455 (B, K, LBV, MO, WAG), forested lower slope of Mount Cameroon, above Batoke; 4°05'N, 9°05'E; alt. 300 m; 24 Apr. 1984 • D.W. Thomas 7372 (MO, P, WAG), Takamanda Forest Reserve; 6°13'N, 9°26'E; alt. 500 m; 30 Apr. 1987 • S.N. Ekema 944 (K, SCA); Ndian, Dikome, Mokoko Forest Reserve; 4°25'11.64'N, 9°03'29.16'E; alt. 120 m; 05 May. 1994 • T.L.P. Couvreur 1027 (MPU, WAG, YA), on trail leading to top of Mt Etinde, after Ekonjo village; 4°04'02.29'N, 9°09'11.15'E; alt. 756 m; 01 Apr. 2016 • T.L.P. Couvreur 978 (WAG, YA), on top of hill, near Small Ekombe village, 3 km after Kumba on road to Ekondo Titi town; 4°37'21.52'N, 9°22'26.62'E; alt. 589 m; 13 Jan. 2016 – Unknown major area • J. Osborne 200 (K, WAG), Littoral Province, Ebo Forest proposed National Park. Ebo Forest Research Station-Bekob drinking stream trail; 4°21'N, 10°25'E; 27 Oct. 2006.
Uvariopsis
Description
Tree ca. 3 m tall, D.B.H unknown; young branches glabrous, old branches glabrous. Petiole ca. 6 mm long, ca. 1.7 mm wide, glabrous. Leaf lamina ca. 160 mm long, ca. 60 mm wide, length:width ratio ca. 2.6, elliptic, coriaceous, base acute to decurrent, apex attenuate, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins ca. 12 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers unknown. Fruits unknown.

Distribution
Endemic to Zambezian Region. Known from only one locality in Uganda: the Budongo Forest Reserve.
Habitat and ecology
Rain forests. Altitude ca. 1051 m asl.
Phenology
Unknown.
Notes
This species resembles Up. congensis in having elliptic leaves with acute to decurrent base and attenuate apex, but it differs in having longer petioles (ca. 6 mm vs. 2–5 mm). Molecular analysis retrieves this species as sister species of the clade formed by Up. congensis and Up. lovettiana. Further collection of this species is needed to provide a more complete description (particularly for the flowers and fruits). This species is known from a single specimen from the Budongo Forest Reserve. It is thus likely to be Critically Endangered CR. However, further exploration of the region is necessary to know whether this species occurs in other localities.
Specimen examined
Uganda – Western Province • ATBP (Africa Tropical Biodiversity Programme) 666 (MO); Masindi District, Budongo Forest Reserve, Nyakafunjo Nature Reserve; 1°43'N, 31°31'E; alt. 1051 m; 26 Jun. 1998.
Uvariopsis
Description
Shrub to tree 4–6 m tall, D.B.H unknown; young branches glabrous, old branches glabrous. Petiole ca. 2 mm long, ca. 2 mm wide, glabrous. Leaf lamina 150–180 mm long, ca. 60 mm wide, length:width ratio ca. 3, elliptic to obovate, coriaceous, base rounded, apex attenuate, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins ca. 13 pairs, brochidodromous to weakly brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers not seen (label on specimen Kyoto University Expedition 1039 says 5 petals). Fruiting pedicel ca. 51 mm long, ca. 1 mm in diameter, pubescent. Monocarps, 1–2, ca. 30 mm long, ca. 15 mm wide, length:width ratio ca. 2, cylindrical to obpyriform, smooth, slightly constricted between the seeds, pubescent, subsessile; stipe 0–1 mm long, ca. 1 mm wide, pubescent.

Distribution
Endemic to Somalia-Masai Region. Only known from the Kigoma region in eastern Tanzania.
Habitat and ecology
Rain forests. Altitude ca. 900 m asl.
Phenology
Fruits collected in December (flowers said to be collected too, but remained not found).
Vernacular names
Tanzania: ‘Ntwelele’ in Tongwe (Kyoto University Expedition 1039).
Notes
This species is very similar to Up. lovettiana having similar elliptic leaf shape, but differs by the leaf base being rounded (vs. acute to decurrent in Up. lovettiana) and in having pubescent monocarps slightly constricted between the seeds (vs. sparsely pubescent to glabrous monocarps, strongly constricted between the seeds). The monocarps are also subsessile (stipe 0–1 mm long, vs. stipitate, stipe 2.5–14 mm long in Up. lovettiana). Confirming the status of new species with molecular analysis is however essential in order to make sure these specimens are not a morphological variation of Up. lovettiana.
Specimens examined
Tanzania – Kigoma • G.S. Bidgood 2783 (DSM, K, P, WAG); Kigoma Rural District, T4. Kigoma Distr.: Kasye Forest; 5°41'S, 29°55'E; alt. 900 m; 18 Mar. 1994 – Kyoto University Expedition 1039 (EA); Kigoma Rural District; 4°45'S, 30°30'E; 24 Dec. 1963.
Uvariopsis
Description
Tree ca. 3.5 m tall, D.B.H unknown; young branches glabrous, old branches glabrous. Petiole ca. 2 mm long, ca. 2 mm wide, glabrous. Leaf lamina ca. 160 mm long, ca. 58 mm wide, length:width ratio ca. 2.8, elliptic, coriaceous, base rounded, apex attenuate, surface above glabrous, surface below glabrous; midrib impressed above, raised below, glabrous above, glabrous below; secondary veins ca. 9 pairs, brochidodromous, impressed above, raised below; tertiary veins reticulate. Flowers only flower bud seen. Flower buds ovoid to conical. Inflorescences axillary, composed of 1 flower. Peduncle inconspicuous. Flower pedicel 20–65 mm long, ca. 1 mm in diameter, glabrate to glabrous. Bracts 1 in the lower half of the pedicel, ca. 1 mm long, ca. 1 mm wide, broadly ovate, sparsely pubescent outside, glabrous inside. Sepals 2, ca. 2 mm long, ca. 3 mm wide. Petals 4, ca. 7 mm long, ca. 4 mm wide, length:width ratio red outside. Fruits unknown.

Distribution
Endemic to Congolia Domain of the Guineo-Congolian Region. Only known from the Kasai-Oriental and Equateur regions in the Democratic Republic of the Congo.
Habitat and ecology
Rain forests.
Phenology
Flowers collected in May and October.
Notes
This species resembles Up. solheidii and Up. noldeae in having elliptic leaves with rounded leaf base. It differs from Up. solheidii in having young branches and petioles completely glabrous (vs. tomentose in Up. solheidii). It differs from Up. noldeae in having flowers borne on young branches and axillary (vs. borne on trunk in Up. noldeae, but see the discussion above about the unreliability of the flower position to discriminate the species), and in having flower pedicels 20–65 mm long (vs. 70–83 mm long in Up. noldeae). This species is known from only two specimens from two different localities distant of more than 700 km from each other in the Congo bassin. It is thus likely to occur in other localities but further exploration of the region is necessary to confirm that.
Specimens examined
Democratic Republic of the Congo – Equateur • C.M. Evrard 6276 (K); Ikela, Yalifake; 0°31'06'N, 22°38'12'E; 06 May. 1959 – Kasai-Oriental • L. Liben 3854 (K); Mwene Ditu, Mwene Ditu, Mwene-Ditu; 7°00'S, 23°27'E; 24 Oct. 1957.
Acknowledgments
The authors acknowledge the ISO 9001 certified IRD i-Trop HPC (member of the South Green Platform, http://www.southgreen.fr) at IRD Montpellier (https://bioinfo.ird.fr/) for providing HPC resources that have contributed to the research results reported within this paper.
In Tanzania, we are grateful to COSTECH for delivering the necessary research permit (permit no. 2019-529-NA-2019-320) and all local authorities for permitting fieldwork. We warmly thank Yusuph Shekilango for the driving and technical support during this fieldwork mission, as well as to all the local guides. In Gabon, we thank the Centre National de la Recherche Scientifique et Technologique (CENAREST), the Agence Nationale des Parcs Nationaux (ANPN), and Prof. Bourobou Bourobou for research permits (AR0020/16 and AR0036/15 [CENAREST] and AE16014 [ANPN]). In Cameroon, fieldwork in Cameroon was undertaken under the “accord cadre de cooperation” between the IRD and Ministere de la Recherche Scientifique et de l’Innovation. We warmly thank Narcisse G. Kamdem for his precious presence on the field. All the specimens were collected with permissions of local authorities and distributed under Material Transfer Agreements.
The visit to the K herbarium was made possible thanks to SYNTHESYS+ grant (GB-TAF-45). We thank Martin Cheek for sending some samples included here. Norbert Holstein is thanked for making the visit of BM possible and fruitful. We are also grateful to B herbarium, EA herbarium and Dr Paul Musili, G herbarium, M herbarium and MO herbarium for the loans of the specimens; to the P herbarium and Dr Thierry Deroin for allowing the visit to the collections. The taxonomic revision work wouldn’t have been possible without MPU herbarium (Montpellier). We also warmly thank the DSM herbarium, Botany Department of Dar es Salaam University, and the Herbier National du Cameroun (YA). We are grateful to all the collectors and co-collectors cited in Suppl. material
We want to warmly thank all the authors of drawings and pictures credited in the figures captions. We are grateful to Zoé Lavocat for the help on the composition of illustrations. We thank Lars Chatrou and Roy Erkens for their thorough evaluation and precious comments during the review process.
Additional information
Conflict of interest
The authors have declared that no competing interests exist.
Ethical statement
No ethical statement was reported.
Funding
This work was supported by the Agence Nationale de la Recherche (AFRODYN Grant Number: ANR-15-CE02-0002-01) and by partial funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program (grant agreement No. 865787).
Author contributions
Conceptualization: LPMJD, TLPC. Data curation: LPMJD, JJW. Formal analysis: LPMJD. Funding acquisition: TLPC, LPMJD. Investigation: MC, FMM, LPMJD. Methodology: TLPC, LPMJD. Project administration: TLPC. Resources: JJW, AG, FMM, MG, MC, CL. Software: LPMJD. Supervision: TLPC. Validation: LPMJD, TLPC. Visualization: LPMJD. Writing – original draft: LPMJD. Writing – review and editing: JJW, BS, TLPC.
Author ORCIDs
Léo-Paul M. J. Dagallier https://orcid.org/0000-0002-3270-1544
Marie Couderc https://orcid.org/0000-0001-7297-155X
Aurélie Grall https://orcid.org/0000-0002-1613-4703
Jan J. Wieringa https://orcid.org/0000-0003-0566-372X
Bonaventure Sonké https://orcid.org/0000-0002-4310-3603
Thomas L. P. Couvreur https://orcid.org/0000-0002-8509-6587
Data availability
All of the data that support the findings of this study are available in the main text, in the Supplementary Information and are deposited at GBIF, the Global Biodiversity Information Facility, available at https://doi.org/10.15468/zdvvkh.
References
- Aiyeloja AA, Bello OA (2006) Ethnobotanical potentials of common herbs in Nigeria: A case study of Enugu State. Educational Research Review 1: 16–22.
- Ajibesin KK, Ekpo BA, Bala DN, Essien EE, Adesanya SA (2008) Ethnobotanical survey of Akwa Ibom State of Nigeria. Journal of Ethnopharmacology 115(3): 387–408. https://doi.org/10.1016/j.jep.2007.10.021
- Bada Amouzoun AAM, Badou RB, Ahamide DYI, Adomou CA (2019) Connaissances ethnobotaniques et conservation de Uvariodendron angustifolium (Engl. & Diels) R. E. Fries (Annonaceae) dans l’îlot forestier Ewe-Adakplame au Sud-Est du Bénin, Afrique de l’Ouest. Revue Ivoirienne des Sciences et Technologie: 328–348.
- Baker EG (1913) Dicotyledons Polypetalae, Annonaceae. Catalogue of the plants collected by Mr. & Mrs. P.A. Talbot in the Oban district, South Nigeria. London, 6 pp. https://www.biodiversitylibrary.org/item/61320
- Bakker FT (2019) Herbarium genomics: plant archival DNA explored. In: Lindqvist C, Rajora OP (Eds) Paleogenomics: Genome-Scale Analysis of Ancient DNA. Population Genomics. Springer International Publishing, Cham, 205–224. https://doi.org/10.1007/13836_2018_40
- Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S, Prjibelski AD, Pyshkin AV, Sirotkin AV, Vyahhi N, Tesler G, Alekseyev MA, Pevzner PA (2012) SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. Journal of Computational Biology 19(5): 455–477. https://doi.org/10.1089/cmb.2012.0021
- Bebber DP, Carine MA, Wood JRI, Wortley AH, Harris DJ, Prance GT, Davidse G, Paige J, Pennington TD, Robson NKB, Scotland RW (2010) Herbaria are a major frontier for species discovery. Proceedings of the National Academy of Sciences of the United States of America 107(51): 22169–22171. https://doi.org/10.1073/pnas.1011841108
- Beentje H (2015) Science comes from collaboration and communication: The Flora of Tropical East Africa as an example. Webbia 70(1): 171–179. https://doi.org/10.1080/00837792.2015.1013244
- Bele MY, Focho DA, Egbe EA, Chuyong BG (2011) Ethnobotanical survey of the uses of Annonaceae around mount Cameroon. African Journal of Plant Science 5: 237–247.
- Bénichou L, Gérard I, Laureys É, Price M (2018) Consortium of European Taxonomic Facilities (CETAF) best practices in electronic publishing in taxonomy. European Journal of Taxonomy (475). https://doi.org/10.5852/ejt.2018.475
- Bentham G (1862) On African Annonaceae. Transactions of the Linnean Society of London 23(3): 463–480. https://doi.org/10.1111/j.1096-3642.1860.tb00144.x
- Binggeli P (2019) Kimboza Forest Reserve. http://www.mikepalmer.co.uk/woodyplantecology/tropical/Kimboza.html
- Bingham MG, Willemen A, Wursten BT, Ballings P, Hyde MA (2021) Flora of Zambia: Home page. https://www.zambiaflora.com/
- Botanic Gardens Conservation International, IUCN SSC Global Tree Specialist Group (2019a) Uvariodendron molundense. The IUCN Red List of Threatened Species 2019: e.T133045155A143272449. https://doi.org/10.2305/IUCN.UK.2019-1.RLTS.T133045155A143272449.en
- Botanic Gardens Conservation International, IUCN SSC Global Tree Specialist Group (2019b) Uvariopsis solheidii. The IUCN Red List of Threatened Species 2019: e.T133045483A143272560. https://doi.org/10.2305/IUCN.UK.2019-1.RLTS.T133045483A143272560.en
- Botermans M, Sosef MSM, Chatrou LW, Couvreur TLP (2011) Revision of the African genus Hexalobus (Annonaceae). Systematic Botany 36(1): 33–48. https://doi.org/10.1600/036364411X553108
- Brée B, Helmstetter AJ, Bethune K, Ghogue J-P, Sonké B, Couvreur TLP (2020) Diversification of African rainforest restricted clades: Piptostigmateae and Annickieae (Annonaceae). Diversity 12(6): e227. https://doi.org/10.3390/d12060227
- Brewer GE, Clarkson JJ, Maurin O, Zuntini AR, Barber V, Bellot S, Biggs N, Cowan RS, Davies NMJ, Dodsworth S, Edwards SL, Eiserhardt WL, Epitawalage N, Frisby S, Grall A, Kersey PJ, Pokorny L, Leitch IJ, Forest F, Baker WJ (2019) Factors affecting targeted sequencing of 353 nuclear genes from herbarium specimens spanning the diversity of Angiosperms. Frontiers in Plant Science 10: e1102. https://doi.org/10.3389/fpls.2019.01102
- Brown JW, Walker JF, Smith SA (2017) Phyx: Phylogenetic tools for unix. Bioinformatics 33(12): 1886–1888. https://doi.org/10.1093/bioinformatics/btx063
- Burgess ND, Clarke GP, Rodgers WA (1998) Coastal forests of eastern Africa: Status, endemism patterns and their potential causes. Biological Journal of the Linnean Society, Linnean Society of London 64(3): 337–367. https://doi.org/10.1111/j.1095-8312.1998.tb00337.x
- Burgess ND, Butynski TM, Cordeiro NJ, Doggart NH, Fjeldså J, Howell KM, Kilahama FB, Loader SP, Lovett JC, Mbilinyi B, Menegon M, Moyer DC, Nashanda E, Perkin A, Rovero F, Stanley WT, Stuart SN (2007) The biological importance of the Eastern Arc Mountains of Tanzania and Kenya. Biological Conservation 134(2): 209–231. https://doi.org/10.1016/j.biocon.2006.08.015
- Burman J (1736) Thesaurus Zeylanicus. Amstelaedamum, 415 pp.
- Chaowasku T, Van der Ham RWJM, Chatrou LW (2013) Integrative systematics supports the establishment of Winitia, a new genus of Annonaceae (Malmeoideae, Miliuseae) allied to Stelechocarpus and Sageraea. Systematics and Biodiversity 11(2): 195–207. https://doi.org/10.1080/14772000.2013.806370
- Chaowasku T, Thomas DC, van der Ham RWJM, Smets EF, Mols JB, Chatrou LW (2014) A plastid DNA phylogeny of tribe Miliuseae: Insights into relationships and character evolution in one of the most recalcitrant major clades of Annonaceae. American Journal of Botany 101(4): 691–709. https://doi.org/10.3732/ajb.1300403
- Chatrou LW, Pirie MD, Erkens RHJ, Couvreur TLP, Neubig KM, Abbott JR, Mols JB, Maas JW, Saunders RMK, Chase MW (2012) A new subfamilial and tribal classification of the pantropical flowering plant family Annonaceae informed by molecular phylogenetics. Botanical Journal of the Linnean Society 169(1): 5–40. https://doi.org/10.1111/j.1095-8339.2012.01235.x
- Cheek M (1989) A new species of Turraea (Meliaceae) from Tanzania. Kew Bulletin 44(3): 465–468. https://doi.org/10.2307/4110366
- Cheek M (2014a) Uvariopsis korupensis. The IUCN Red List of Threatened Species 2014: e.T200683A2678951. https://doi.org/10.2305/IUCN.UK.2014-3.RLTS.T200683A2678951.en
- Cheek M (2014b) Uvariopsis submontana. The IUCN Red List of Threatened Species 2014: e.T45421A3001680. https://doi.org/10.2305/IUCN.UK.2014-3.RLTS.T45421A3001680.en
- Cheek M (2014c) Uvariopsis vanderystii. The IUCN Red List of Threatened Species 2014: e.T45422A3001751. https://doi.org/10.2305/IUCN.UK.2014-3.RLTS.T45422A3001751.en
- Cheek M, Cable S (2000) Uvariodendron fuscum. The IUCN Red List of Threatened Species 2000: e.T39476A10240275. https://doi.org/10.2305/IUCN.UK.2000.RLTS.T39476A10240275.en
- Cheek M, Dorr LJ, Beentje HJ, Ghazanfar SA (2007) Flora of tropical East Africa – Sterculiaceae. publ. on behalf of the East African governments by Royal Botanic Gardens, Kew.
- Cheek M, Luke Q, Matimele H, Banze A, Lawrence P (2019) Cola species of the limestone forests of Africa, with a new, endangered species, Cola cheringoma (Sterculiaceae), from Cheringoma, Mozambique. Kew Bulletin 74(4): 1–52. https://doi.org/10.1007/s12225-019-9840-3
- Cheek M, Luke WRQ, Gosline G (2022) Lukea gen. nov. (Monodoreae-Annonaceae) with two new threatened species of shrub from the forests of the Udzungwas, Tanzania and Kaya Ribe, Kenya. Kew Bulletin 77(3): 647–664. https://doi.org/10.1007/s12225-022-10039-y
- Chevalier A (1920) Exploration Botanique de l’Afrique Occidentale Française. Paul Lechevallier, Paris.
- Cosiaux A, Couvreur TLP, Erkens RHJ (2019a) Uvariodendron calophyllum. The IUCN Red List of Threatened Species 2019: e.T133045070A133045322. https://doi.org/10.2305/IUCN.UK.2019-3.RLTS.T133045070A133045322.en
- Cosiaux A, Couvreur TLP, Erkens RHJ (2019b) Uvariopsis bakeriana. The IUCN Red List of Threatened Species 2019: e.T133045364A133046152. https://doi.org/10.2305/IUCN.UK.2019-3.RLTS.T133045364A133046152.en
- Cosiaux A, Couvreur TLP, Erkens RHJ (2019c) Uvariopsis citrata. The IUCN Red List of Threatened Species 2019: e.T133045382A133046157. https://doi.org/10.2305/IUCN.UK.2019-3.RLTS.T133045382A133046157.en
- Cosiaux A, Couvreur TLP, Erkens RHJ (2019d) Uvariopsis noldeae. The IUCN Red List of Threatened Species 2019: e.T137012111A137048277. https://doi.org/10.2305/IUCN.UK.2019-3.RLTS.T137012111A137048277.en
- Couvreur TLP (2009) Monograph of the syncarpous African genera Isolona and Monodora (Annonaceae). Systematic Botany Monographs 87: 1–150.
- Couvreur TLP (2014) Revision of the African genus Uvariastrum (Annonaceae). PhytoKeys 33: 1–40. https://doi.org/10.3897/phytokeys.33.5907
- Couvreur TLP, Luke WRQ (2010) A new species of Uvariopsis (Annonaceae), endemic to the Eastern Arc Mountains of Tanzania. Blumea 55(1): 68–72. https://doi.org/10.3767/000651910X499196
- Couvreur TLP, Niangadouma R (2016) New species of Uvariopsis (Annonaceae) and Laccosperma (Arecaceae/Palmae) from Monts de Cristal, Gabon. PhytoKeys 68: 1–8. https://doi.org/10.3897/phytokeys.68.9576
- Couvreur TLP, Chatrou LW, Sosef MSM, Richardson JE (2008a) Molecular phylogenetics reveal multiple Tertiary vicariance origins of the African rain forest trees. BMC Biology 6(1): 1–54. https://doi.org/10.1186/1741-7007-6-54
- Couvreur TLP, Richardson JE, Sosef MSM, Erkens RHJ, Chatrou LW (2008b) Evolution of syncarpy and other morphological characters in African Annonaceae: A posterior mapping approach. Molecular Phylogenetics and Evolution 47(1): 302–318. https://doi.org/10.1016/j.ympev.2008.01.018
- Couvreur TLP, van der Ham RWJM, Mbele YM, Mbago FM, Johnson DM (2009) Molecular and morphological characterization of a new monotypic genus of Annonaceae, Mwasumbia, from Tanzania. Systematic Botany 34(2): 266–276. https://doi.org/10.1600/036364409788606398
- Couvreur TLP, Helmstetter AJ, Koenen EJM, Bethune K, Brandão RD, Little SA, Sauquet H, Erkens RHJ (2019) Phylogenomics of the major tropical plant family Annonaceae using targeted enrichment of nuclear genes. Frontiers in Plant Science 9: e1941. https://doi.org/10.3389/fpls.2018.01941
- Couvreur TLP, Dagallier L-PMJ, Crozier F, Ghogue J-P, Hoekstra PH, Kamdem NG, Johnson DM, Murray NA, Sonké B (2022) Flora of Cameroon – Annonaceae (Vol. 45). PhytoKeys 207: 1–532. https://doi.org/10.3897/phytokeys.207.61432
- Dagallier LMJ, Janssens SB, Dauby G, Blach‐Overgaard A, Mackinder BA, Droissart V, Svenning J, Sosef MSM, Stévart T, Harris DJ, Sonké B, Wieringa JJ, Hardy OJ, Couvreur TLP (2020) Cradles and museums of generic plant diversity across tropical Africa. The New Phytologist 225(5): 2196–2213. https://doi.org/10.1111/nph.16293
- Dagallier L-PMJ, Mbago FM, Luke WRQ, Couvreur TLP (2021) Three new species of Uvariodendron (Annonaceae) from coastal East Africa in Kenya and Tanzania. PhytoKeys 174: 107–126. https://doi.org/10.3897/phytokeys.174.61630
- Dauby G, Stévart T, Droissart V, Cosiaux A, Deblauwe V, Simo-Droissart M, Sosef MSM, Lowry PP II, Schatz GE, Gereau RE, Couvreur TLP (2017) ConR: An R package to assist large-scale multispecies preliminary conservation assessments using distribution data. Ecology and Evolution 7(24): 11292–11303. https://doi.org/10.1002/ece3.3704
- De Wildeman E (1909) Flore du Bas et du Moyen Congo. Annales du Musée du Congo Belge, Botanique, Série 5 3: 1–147.
- Deroin T (1985) Contribution à la morphologie comparée du gynécée des Annonaceae – Monodoroideae. Bulletin du Muséum National d’Histoire Naturelle, Section B, Adansonia 7: 167–176.
- Diels L (1907) Anonaceae africanae. Botanische Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie 39: 469–486. https://www.biodiversitylibrary.org/item/137011
- Diels L (1915) Anonaceae africanae III. In: Engler A (Ed. ) Botanische Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie 53: 434–448.
- Diels L (1936) Annonaceae, Neue und seltene Arten aus Ostafrika. Notizblatt des Botanischen Gartens und Museums zu Berlin-Dahlem 13: 165–272.
- Doyle JA, Le Thomas A (2012) Evolution and phylogenetic significance of pollen in Annonaceae. Botanical Journal of the Linnean Society 169(1): 190–221. https://doi.org/10.1111/j.1095-8339.2012.01241.x
- Doyle JA, Sauquet H, Scharaschkin T, Le Thomas A (2004) Phylogeny, molecular and fossil dating, and biogeographic history of Annonaceae and Myristicaceae (Magnoliales). International Journal of Plant Sciences 165(S4): S55–S67. https://doi.org/10.1086/421068
- Droissart V, Dauby G, Hardy OJ, Deblauwe V, Harris DJ, Janssens S, Mackinder BA, Blach-Overgaard A, Sonké B, Sosef MSM, Stévart T, Svenning J-C, Wieringa JJ, Couvreur TLP (2018) Beyond trees: Biogeographical regionalization of tropical Africa. Journal of Biogeography 45(5): 1153–1167. https://doi.org/10.1111/jbi.13190
- Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project (2009a) Uvariodendron gorgonis. The IUCN Red List of Threatened Species 2009: e.T32242A9690711. https://doi.org/10.2305/IUCN.UK.2009-2.RLTS.T32242A9690711.en
- Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project (2009b) Uvariodendron kirkii. The IUCN Red List of Threatened Species 2009: e.T35724A9953716. https://doi.org/10.2305/IUCN.UK.2009-2.RLTS.T35724A9953716.en
- Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project (2009c) Uvariodendron pycnophyllum. The IUCN Red List of Threatened Species 2009: e.T32861A9729115. https://doi.org/10.2305/IUCN.UK.2009-2.RLTS.T32861A9729115.en
- Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project (2009d) Uvariodendron usambarense. The IUCN Red List of Threatened Species 2009: e.T32862A9729743. https://doi.org/10.2305/IUCN.UK.2009-2.RLTS.T32862A9729743.en
- Eastern Arc Mountains & Coastal Forests CEPF Plant Assessment Project (2009e) Uvariopsis bisexualis. The IUCN Red List of Threatened Species 2009: e.T35389A9925787. https://doi.org/10.2305/IUCN.UK.2009-2.RLTS.T35389A9925787.en
- Engler A, Diels L (1899) Annonaceae. Notizblatt des Königl. Botanischen Gartens und Museums zu Berlin. 2: 292–308.
- Engler A, Diels L (1901) Monographieen afrikanischer Pflanzen-Familien und -Gattungen – Annonaceae. W. Engelmann, Leipzig, 162 pp. https://www.biodiversitylibrary.org/item/113590
- Erkens RHJ, Chatrou LW, Couvreur TLP (2012) Radiations and key innovations in an early branching angiosperm lineage (Annonaceae; Magnoliales). Botanical Journal of the Linnean Society 169(1): 117–134. https://doi.org/10.1111/j.1095-8339.2012.01223.x
- Erkens RHJ, Chatrou LW, Chaowasku T, Westra LYT, Maas JW, Maas PJM (2014) A decade of uncertainty: Resolving the phylogenetic position of Diclinanona (Annonaceae), including taxonomic notes and a key to the species. Taxon 63(6): 1244–1252. https://doi.org/10.12705/636.34
- Erkens RHJ, Oosterhof J, Westra LYT, Maas PJM (2017) Revisions of Ruizodendron and Pseudephedranthus (Annonaceae) including a new species and an overview of most up-to-date revisions of Neotropical Annonaceae genera. PhytoKeys 86: 75–96. https://doi.org/10.3897/phytokeys.86.13773
- Exell AW (1926) Mr. John Gossweiler’s plants from Angola and Portuguese Congo. Journal of Botany 64: Suppl. 1: Polypetalae: 1–24.
- Exell AW, Mendonça FA (1937) 1 Conspectus florae Angolensis. Instituto Botanico de Coimbra, Coimbra, 422 pp.
- Exell AW, Mendonça F de A (1951) Novidades da Flora de Angola. Departamento de Botânica-FCTUC, Coimbra, 101 pp.
- Exell AW, Wild H (1960) Flora zambesiaca (Vol. 1, Part I). Crown Agents, London. https://www.cabdirect.org/cabdirect/abstract/19600303022
- Fjeldså J, Lovett JC (1997) Geographical patterns of old and young species in African forest biota: The significance of specific montane areas as evolutionary centres. Biodiversity and Conservation 6(3): 325–346. https://doi.org/10.1023/A:1018356506390
- Fries RE (1930) Revision der Arten einiger Anonaceen-Gattungen. Acta Horti Bergiani 10: 1–128.
- Fries RE (1953) Verstreute Beobachtungen hinsichtlich der Familie Annonaceae. Arkiv för botanik ser.2 3: 35–42.
- Gautier-Hion A, Duplantier J-M, Quris R, Feer F, Sourd C, Decoux J-P, Dubost G, Emmons L, Erard C, Hecketsweiler P, Moungazi A, Roussilhon C, Thiollay J-M (1985) Fruit characters as a basis of fruit choice and seed dispersal in a tropical forest vertebrate community. Oecologia 65(3): 324–337. https://doi.org/10.1007/BF00378906
- Gereau RE, Kenfack D (2000) Le genre Uvariopsis (Annonaceae) en Afrique tropicale, avec la description d’une espèce nouvelle du Cameroun. Adansonia 22: 39–43.
- Gosline G, Marshall AR, Larridon I (2019) Revision and new species of the African genus Mischogyne (Annonaceae). Kew Bulletin 74(2): 1–28. https://doi.org/10.1007/s12225-019-9804-7
- Gosline G, Cheek M, Onana JM, Tchatchouang EN, van der Burgt XM, MacKinnon L, Dagallier L-PMJ (2022) Uvariopsis dicaprio (Annonaceae) a new tree species with notes on its pollination biology, and the Critically Endangered narrowly endemic plant species of the Ebo Forest, Cameroon. PeerJ 9: e12614. https://doi.org/10.7717/peerj.12614
- Gottsberger G, Webber AC (2018) Nutritious tissue in petals of Annonaceae and its function in pollination by scarab beetles. Acta Botanica Brasílica 32(2): 279–286. https://doi.org/10.1590/0102-33062017abb0356
- Gottsberger G, Meinke S, Porembski S (2011) First records of flower biology and pollination in African Annonaceae: Isolona, Piptostigma, Uvariodendron, Monodora and Uvariopsis. Flora – Morphology, Distribution. Flora 206(5): 498–510. https://doi.org/10.1016/j.flora.2010.08.005
- Guillocheau F, Simon B, Baby G, Bessin P, Robin C, Dauteuil O (2018) Planation surfaces as a record of mantle dynamics: The case example of Africa. Gondwana Research 53: 82–98. https://doi.org/10.1016/j.gr.2017.05.015
- Guo X, Tang CC, Thomas DC, Couvreur TLP, Saunders RMK (2017a) A mega-phylogeny of the Annonaceae: Taxonomic placement of five enigmatic genera and support for a new tribe, Phoenicantheae. Scientific Reports 7(1): e7323. https://doi.org/10.1038/s41598-017-07252-2
- Guo X, Hoekstra PH, Tang CC, Thomas DC, Wieringa JJ, Chatrou LW, Saunders RMK (2017b) Cutting up the climbers: Evidence for extensive polyphyly in Friesodielsia (Annonaceae) necessitates generic realignment across the tribe Uvarieae. Taxon 66(1): 3–19. https://doi.org/10.12705/661.1
- Guo X, Thomas DC, Saunders RMK (2018) Gene tree discordance and coalescent methods support ancient intergeneric hybridisation between Dasymaschalon and Friesodielsia (Annonaceae). Molecular Phylogenetics and Evolution 127: 14–29. https://doi.org/10.1016/j.ympev.2018.04.009
- Hall JB, Rodgers WA (1986) Pole cutting pressure in Tanzanian forest. Forest Ecology and Management 14(2): 133–140. https://doi.org/10.1016/0378-1127(86)90098-8
- Hardy OJ, Born C, Budde K, Daïnou K, Dauby G, Duminil J, Ewédjé E-EBK, Gomez C, Heuertz M, Koffi GK, Lowe AJ, Micheneau C, Ndiade-Bourobou D, Piñeiro R, Poncet V (2013) Comparative phylogeography of African rain forest trees: A review of genetic signatures of vegetation history in the Guineo-Congolian region. Comptes Rendus Geoscience 345(7–8): 284–296. https://doi.org/10.1016/j.crte.2013.05.001
- Harris JG, Harris MW (2001) Plant Identification Terminology: An Illustrated Glossary. Spring Lake publishing, Spring Lake, Utah, Etats-Unis d’Amérique, 206 pp.
- Harvey-Brown Y (2019a) Uvariopsis congensis. The IUCN Red List of Threatened Species 2019: e.T133045390A149069038. https://doi.org/10.2305/IUCN.UK.2019-2.RLTS.T133045390A149069038.en
- Harvey-Brown Y (2019b) Uvariopsis congolana. The IUCN Red List of Threatened Species 2019: e.T133045403A149068858. https://doi.org/10.2305/IUCN.UK.2019-2.RLTS.T133045403A149068858.en
- Harvey-Brown Y (2019c) Uvariopsis guineensis. The IUCN Red List of Threatened Species 2019: e.T85730032A149068563. https://doi.org/10.2305/IUCN.UK.2019-2.RLTS.T85730032A149068563.en
- Hawthorne W (1998) Uvariodendron occidentale. The IUCN Red List of Threatened Species 1998: e.T34826A9892349. https://doi.org/10.2305/IUCN.UK.1998.RLTS.T34826A9892349.en
- Hawthorne WD, Jongkind CCH (2006) Woody plants of Western African forests, A guide to the forest trees, shrubs and lianes from Senegal to Ghana. Royal Botanic Gardens, Kew, Richmond, Surrey, 1040 pp.
- Helmstetter AJ, Béthune K, Kamdem NG, Sonké B, Couvreur TLP (2020a) Individualistic evolutionary responses of Central African rain forest plants to Pleistocene climatic fluctuations. Proceedings of the National Academy of Sciences of the United States of America 117(51): 32509–32518. https://doi.org/10.1073/pnas.2001018117
- Helmstetter AJ, Kamga SM, Bethune K, Lautenschläger T, Zizka A, Bacon CD, Wieringa JJ, Stauffer F, Antonelli A, Sonké B, Couvreur TLP (2020b) Unraveling the phylogenomic relationships of the most diverse African palm genus Raphia (Calamoideae, Arecaceae). Plants 9(4): e549. https://doi.org/10.3390/plants9040549
- Hickey LJ, Ellis B, Daly DC, Johnson KR, Mitchell JD, Wilf P, Wing SL (1999) Manual of leaf architecture: morphological description and categorization of dicotyledonous and net-veined monocotyledonous angiosperms. Leaf Architecture Working Group, Washington, 65 pp.
- Hoekstra PH, Wieringa JJ, Maas PJM, Chatrou LW (2021) Revision of the African species of Monanthotaxis (Annonaceae). Blumea 66(2): 107–221. https://doi.org/10.3767/blumea.2021.66.02.01
- Hutchinson J, Dalziel JM (1927a) Flora of West Tropical Africa (1st ed.). Crown Agents for the Colonies, London, 125 pp.
- Hutchinson J, Dalziel JM (1927b) Tropical African Plants: I. Bulletin of Miscellaneous Information 1927: 150–157. https://doi.org/10.2307/4107589
- Hyde MA, Wursten BT, Ballings P, Coates Palgrave M (2021) Flora of Mozambique. https://www.mozambiqueflora.com/ [July 26, 2021]
- Iseghohi SO (2015) A review of the uses and medicinal properties of Dennettia tripetala (Pepperfruit). Medical Sciences: Open Access Journal 3(4): 104–111. https://doi.org/10.3390/medsci3040104
- IUCN (2012) IUCN Red List Categories and Criteria: Version 3.1. (2nd Edn.). IUCN, Gland, Switzerland and Cambridge, 32 pp.
- Jiofack T, Fokunang C, Guedje N, Kemeuze V, Fongnzossie E, Nkongmeneck BA, Mapongmetsem PM, Tsabang N (2009) Ethnobotanical uses of some plants of two ethnoecological regions of Cameroon. African Journal of Pharmacy and Pharmacology 3: 664–684. https://academicjournals.org/journal/AJPP/article-abstract/15BDBA338011
- Johnson DM (2003) Phylogenetic significance of spiral and distichous architecture in the Annonaceae. Systematic Botany 28: 503–512.
- Johnson MG, Gardner EM, Liu Y, Medina R, Goffinet B, Shaw AJ, Zerega NJC, Wickett NJ (2016) HybPiper: Extracting coding sequence and introns for phylogenetics from high-throughput sequencing reads using target enrichment. Applications in Plant Sciences 4(7): e1600016. https://doi.org/10.3732/apps.1600016
- Kagoro-Rugunda G, Hashimoto C (2015) Fruit phenology of tree species and chimpanzees’ choice of consumption in Kalinzu Forest Reserve, Uganda. Open Journal of Ecology 5(10): 477–490. https://doi.org/10.4236/oje.2015.510039
- Kaingu CK, Oduma JA, Mbaria J, Kiama SG (2017) Effects of Croton menyharthii and Uvariodendron kirkii extracts on ovarian corpora lutea and reproductive hormones. Discovery Phytomedicine 4(2): 21–25. https://doi.org/10.15562/phytomedicine.2017.41
- Katoh K, Standley DM (2013) MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Molecular Biology and Evolution 30(4): 772–780. https://doi.org/10.1093/molbev/mst010
- Keay RWJ (1952) Revision of the “Flora of West Tropical Africa”: I. Kew Bulletin 7(2): 149–165. https://doi.org/10.2307/4109256
- Keay RWJ (1953) Revision of the “Flora of West Tropical Africa”: III. Kew Bulletin 8(1): 69–73. https://doi.org/10.2307/4117161
- Kenfack D, Gosline G, Gereau RE, Schatz GE (2003) The genus Uvariopsis (Annonaceae) in tropical Africa, with a recombination and one new species from Cameroon. Novon 13(4): 443–449. https://doi.org/10.2307/3393377
- Kuntze O (1891) 1 Revisio Generum Plantarum. Charles Klincksieck, Paris, 533 pp. https://www.biodiversitylibrary.org/item/7555
- Kuntze O (1903) Inkorrekte Benennungen neuer Species in Englers Notizblatt des Könrigl. botanischen Gartens und Museums zu Berlin. Deutsche botanische Monatsschrift 21: 172–174.
- Lambert JE (2011) Primate seed dispersers as umbrella species: A case study from Kibale National Park, Uganda, with implications for Afrotropical forest conservation. American Journal of Primatology 73(1): 9–24. https://doi.org/10.1002/ajp.20879
- Le Thomas A (1965) Notes sur quelques Annonaceae Ouest-Africaines. Adansonia 5: 443–454.
- Le Thomas A (1967) A propos de l’Uvariodendron mirabile R. E. Fries. Adansonia VII: 249–253.
- Le Thomas A (1969) Flore du Gabon 16 – Annonacées. Muséum National d’Histoire Naturelle, Paris, 371 pp.
- Li H, Durbin R (2009) Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25(14): 1754–1760. https://doi.org/10.1093/bioinformatics/btp324
- Lopes J de C, Chatrou LW, Mello-Silva R (2014) Ephedranthus dimerus (Annonaceae), a new species from the Atlantic Forest of Brazil, with a key to the species of Ephedranthus. Brittonia 66(1): 70–74. https://doi.org/10.1007/s12228-013-9311-3
- Lopes JC, Chatrou LW, Mello-Silva R, Rudall PJ, Sajo MG (2018) Phylogenomics and evolution of floral traits in the Neotropical tribe Malmeeae (Annonaceae). Molecular Phylogenetics and Evolution 118: 379–391. https://doi.org/10.1016/j.ympev.2017.10.020
- Luke WRQ, Musili P, Barasa J, Mathenge J (2018) Uvariodendron anisatum. The IUCN Red List of Threatened Species 2018: e.T32241A128399203. https://doi.org/10.2305/IUCN.UK.2018-2.RLTS.T32241A128399203.en [September 21, 2022]
- Maddison WP (1997) Gene trees in species trees. Systematic Biology 46(3): 523–536. https://doi.org/10.1093/sysbio/46.3.523
- Martin M (2011) Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. Journal 17(1): 10–12. https://doi.org/10.14806/ej.17.1.200
- Meade CV, Parnell JAN (2018) A revised taxonomy for Uvaria (Annonaceae) in continental Asia. Australian Systematic Botany 31(4): 311–356. https://doi.org/10.1071/SB17051
- Mertens JEJ, Tropek R, Dzekashu FF, Maicher V, Fokam EB, Janeček Š (2018) Communities of flower visitors of Uvariopsis dioica (Annonaceae) in lowland forests of Mt. Cameroon, with notes on its potential pollinators. African Journal of Ecology 56(1): 146–152. https://doi.org/10.1111/aje.12429
- Meyer BS, Matschiner M, Salzburger W (2017) Disentangling incomplete lineage sorting and introgression to refine species-tree estimates for Lake Tanganyika Cichlid fishes. Systematic Biology 66: 531–550. https://doi.org/10.1093/sysbio/syw069
- Migliore J, Kaymak E, Mariac C, Couvreur TLP, Lissambou B-J, Piñeiro R, Hardy OJ (2019) Pre-Pleistocene origin of phylogeographical breaks in African rain forest trees: New insights from Greenwayodendron (Annonaceae) phylogenomics. Journal of Biogeography 46: 212–223. https://doi.org/10.1111/jbi.13476
- Mildbraed GWJ, Diels L (1915) Anonaceae Africanae III. Botanische Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie 53: 434–448.
- Morales‐Briones DF, Liston A, Tank DC (2018) Phylogenomic analyses reveal a deep history of hybridization and polyploidy in the Neotropical genus Lachemilla (Rosaceae). The New Phytologist 218(4): 1668–1684. https://doi.org/10.1111/nph.15099
- Muhammed D, Adebiyi YH, Odey BO, Alawode RA, Lawal A, Okunlola BM, Ibrahim J, Berinyuy EB (2021) Dennettia tripetala (Pepper Fruit), a review of its ethno-medicinal use, phyto-constituents, and biological properties. GSC Advanced Research and Reviews 6: 035–043. https://doi.org/10.30574/gscarr.2021.6.3.0024
- Naciri Y, Linder HP (2015) Species delimitation and relationships: The dance of the seven veils. Taxon 64(1): 3–16. https://doi.org/10.12705/641.24
- Obua J, Agea JG, Ogwal JJ (2010) Status of forests in Uganda. African Journal of Ecology 48(4): 853–859. https://doi.org/10.1111/j.1365-2028.2010.01217.x
- Olatokunbo IS, Arowosegbe S, Kayode J, Oyedeji A (2018) Ethnobotanical survey of plant species utilised as spices among the indigenous people of Bayelsa State, Nigeria. Brazilian Journal of Biological Sciences 5(10): 461–469. https://doi.org/10.21472/bjbs.051022
- Omage SO, Orhue NEJ, Omage K (2019) Evaluation of the phytochemical content, in vitro antioxidant capacity, biochemical and histological effects of Dennettia tripetala fruits in healthy rats. Food Science & Nutrition 7(1): 65–75. https://doi.org/10.1002/fsn3.792
- Ortiz-Rodriguez AE, Ornelas JF, Ruiz-Sanchez E (2018) A jungle tale: Molecular phylogeny and divergence time estimates of the Desmopsis – Stenanona clade (Annonaceae) in Mesoamerica. Molecular Phylogenetics and Evolution 122: 80–94. https://doi.org/10.1016/j.ympev.2018.01.021
- Paiva JAR (1966) Revisão das Annonaceae de Angola. Memórias da Sociedade Broteriana 19: 5–124.
- Parins-Fukuchi C, Stull GW, Smith SA (2021) Phylogenomic conflict coincides with rapid morphological innovation. Proceedings of the National Academy of Sciences of the United States of America 118(19): e2023058118. https://doi.org/10.1073/pnas.2023058118
- Patrick C (2008) An impact of invasive species Cedrela odorata L. on native plant species composition and diversity in Kimboza forest reserve; Morogoro, Tanzania. Thesis. University of Dar es Salaam.
- Payne WW (1978) A glossary of plant hair terminology. Brittonia 30(2): 239–255. https://doi.org/10.2307/2806659
- Peguy T (1998) Uvariodendron connivens. The IUCN Red List of Threatened Species 1998: e.T34766A9883333. https://doi.org/10.2305/IUCN.UK.1998.RLTS.T34766A9883333.en
- Pellegrin F (1920) Plantae Letestuanae Novae ou Plantes Nouvelles récoltées par M. Le Testu de 1907 à 1919 dans le Mayombe Congolais. Bulletin du Muséum National d’Histoire Naturelle 26: 654–659.
- Pellegrin F (1948) Les Annonacées du Gabon. Bulletin de la Société Botanique de France 95(3–4): 136–141. https://doi.org/10.1080/00378941.1948.10834679
- Pennington RT, Lavin M (2016) The contrasting nature of woody plant species in different neotropical forest biomes reflects differences in ecological stability. The New Phytologist 210(1): 25–37. https://doi.org/10.1111/nph.13724
- Pirie MD, Chatrou LW, Maas PJM (2018) A taxonomic revision of the Neotropical genus Cremastosperma (Annonaceae), including five new species. PhytoKeys 112: 1–141. https://doi.org/10.3897/phytokeys.112.24897
- Plumptre AJ, Ayebare S, Behangana M, Forrest TG, Hatanga P, Kabuye C, Kirunda B, Kityo R, Mugabe H, Namaganda M, Nampindo S, Nangendo G, Nkuutu DN, Pomeroy D, Tushabe H, Prinsloo S (2019) Conservation of vertebrates and plants in Uganda: Identifying Key Biodiversity Areas and other sites of national importance. Conservation Science and Practice 1(2): e7. https://doi.org/10.1002/csp2.7
- R Core Team (2022) R: a language and environment for statistical computing. Austria. https://www.R-project.org
- Rafiu B, Sonibare MA (2016) Ethnobotanical survey of tree species used for wound healing in Ibadan, southwest Nigeria. Nigerian Journal of Natural Products and Medicine 20(0): 96–104. https://doi.org/10.4314/njnpm.v20i0.16
- Rahbek C, Borregaard MK, Antonelli A, Colwell RK, Holt BG, Nogues-Bravo D, Rasmussen CMØ, Richardson K, Rosing MT, Whittaker RJ, Fjeldså J (2019) Building mountain biodiversity: Geological and evolutionary processes. Science 365(6458): 1114–1119. https://doi.org/10.1126/science.aax0151
- Reginato M (2016) monographaR: An R package to facilitate the production of plant taxonomic monographs. Brittonia 68(2): 212–216. https://doi.org/10.1007/s12228-015-9407-z
- Richardson JE, Chatrou LW, Mols JB, Erkens RHJ, Pirie MD (2004) Historical biogeography of two cosmopolitan families of flowering plants: Annonaceae and Rhamnaceae. Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences 359(1450): 1495–1508. https://doi.org/10.1098/rstb.2004.1537
- Robson NKB (1960) Annonaceae. Flora zambesiaca (Vol. 1, Part I). Crown Agents, London, 104–149. https://www.cabdirect.org/cabdirect/abstract/19600303022
- Robyns W, Ghesquière J (1933a) Essai de revision des genres Uvariopsis Engl. & Diels et Tetrastemma Diels (Annonacées). Annales de la Société scientifique de Bruxelles Sér. B: 312–322.
- Robyns W, Ghesquière J (1933b) Quelques Annonacées nouvelles du Congo Belge. Annales de la Société scientifique de Bruxelles Sér. B: 62–69.
- Rogers ME, Abernethy K, Bermejo M, Cipolletta C, Doran D, Mcfarland K, Nishihara T, Remis M, Tutin CEG (2004) Western gorilla diet: A synthesis from six sites. American Journal of Primatology 64(2): 173–192. https://doi.org/10.1002/ajp.20071
- Rodgers WA, Hall JB, Mwasumbi LB, Griffiths CJ, Vollesen K (1983) The Conservation Values and Status of Kimboza Forest Reserve, Tanzania. World Wide Fund for Nature, 84 pp.
- Saunders RMK (2010) Floral evolution in the Annonaceae: Hypotheses of homeotic mutations and functional convergence. Biological Reviews of the Cambridge Philosophical Society 85: 571–591. https://doi.org/10.1111/j.1469-185X.2009.00116.x
- Scharff N (1992) The linyphiid fauna of eastern Africa (Araneae: Linyphiidae)-distribution patterns, diversity and endemism. Biological Journal of the Linnean Society. Linnean Society of London 45(2): 117–154. https://doi.org/10.1111/j.1095-8312.1992.tb00635.x
- Schrempf D, Szöllősi G (2020) The Sources of Phylogenetic Conflicts. In: Phylogenetics in the Genomic Era. No commercial publisher | Authors open access book, 247–269. https://hal.archives-ouvertes.fr/hal-02535070
- Shen X-X, Steenwyk JL, Rokas A (2021) Dissecting incongruence between concatenation- and quartet-based approaches in phylogenomic data. Systematic Biology 70(5): 997–1014. https://doi.org/10.1093/sysbio/syab011
- Sigovini M, Keppel E, Tagliapietra D (2016) Open Nomenclature in the biodiversity era. Methods in Ecology and Evolution 7(10): 1217–1225. https://doi.org/10.1111/2041-210X.12594
- Simon R, Spooner DM (2013) A new R package, exsic, to assist taxonomists in creating indices. Applications in Plant Sciences 1(6): e1300024. https://doi.org/10.3732/apps.1300024
- Sonké B, Couvreur TLP (2014) Tree diversity of the Dja Faunal Reserve, southeastern Cameroon. Biodiversity Data Journal 1049: e1049. https://doi.org/10.3897/BDJ.2.e1049
- Sosef MSM, Dauby G, Blach-Overgaard A, van der Burgt X, Catarino L, Damen T, Deblauwe V, Dessein S, Dransfield J, Droissart V, Duarte MC, Engledow H, Fadeur G, Figueira R, Gereau RE, Hardy OJ, Harris DJ, de Heij J, Janssens S, Klomberg Y, Ley AC, Mackinder BA, Meerts P, van de Poel JL, Sonké B, Stévart T, Stoffelen P, Svenning J-C, Sepulchre P, Zaiss R, Wieringa JJ, Couvreur TLP (2017) Exploring the floristic diversity of tropical Africa. BMC Biology 15(1): 1–15. https://doi.org/10.1186/s12915-017-0356-8
- Stamatakis A (2014) RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30(9): 1312–1313. https://doi.org/10.1093/bioinformatics/btu033
- Synnott TJ (1985) A checklist of the flora of Budongo Forest Reserve, Uganda, with notes on ecology and phenology. Commonwealth Forestry Institute, University of Oxford, UK. https://ora.ox.ac.uk/objects/uuid:a2dd42bc-787c-4b88-931f-ad51c2f26528
- Systematics Association Committee for Descriptive Biological Terminology (1962) II. Terminology of Simple Symmetrical Plane Shapes (Chart 1). Taxon 11(5): 145–156. https://doi.org/10.2307/1216718
- Talavera G, Castresana J (2007) Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Systematic Biology 56(4): 564–577. https://doi.org/10.1080/10635150701472164
- Tang CC, Thomas DC, Saunders RMK (2015) Molecular phylogenetics of the species-rich angiosperm genus Goniothalamus (Annonaceae) inferred from nine chloroplast DNA regions: Synapomorphies and putative correlated evolutionary changes in fruit and seed morphology. Molecular Phylogenetics and Evolution 92: 124–139. https://doi.org/10.1016/j.ympev.2015.06.016
- Texier N, Stévart T (2021) Uvariopsis zenkeri. The IUCN Red List of Threatened Species 2021: e.T133046133A133046192. https://doi.org/10.2305/IUCN.UK.2021-3.RLTS.T133046133A133046192.en
- Thiers B (2016) Index Herbariorum: A global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. https://doi.org/10.1007/s12228-016-9423-7
- Thomas DC, Surveswaran S, Xue B, Sankowsky G, Mols JB, Keßler PJA, Saunders RMK (2012) Molecular phylogenetics and historical biogeography of the Meiogyne–Fitzalania clade (Annonaceae): Generic paraphyly and late Miocene–Pliocene diversification in Australasia and the Pacific. Taxon 61(3): 559–575. https://doi.org/10.1002/tax.613006
- Turland NJ, Wiersema JH, Barrie FR, Greuter W, Hawksworth DL, Herendeen PS, Knapp S, Kusber W-H, Li D-Z, Marhold K, May TW, McNeill J, Monro AM, Prado J, Price MJ, Smith GF (2018) International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Koeltz Botanical Books. https://doi.org/10.12705/Code.2018
- Turner IM (2018) Annonaceae of the Asia-Pacific region: Names, types and distributions. Gardens’ Bulletin 70(2): 409–744. https://doi.org/10.26492/gbs70(2).2018-11
- van Heusden ECH (1992) Flowers of Annonaceae: Morphology, classification, and evolution. Blumea 7(Supplement): 1–218.
- Verdcourt B (1955) Notes from the East African Herbarium: III. Kew Bulletin 10(4): 595–609. https://doi.org/10.2307/4113772
- Verdcourt B (1960) Notes from the East African Herbarium: XI. Kew Bulletin 14(3): 345–352. https://doi.org/10.2307/4114750
- Verdcourt B (1969) The genus Uvariodendron (Engl. & Diels) R. E. Fries (Annonaceae) in East Africa. Kew Bulletin 23(3): 511–521. https://doi.org/10.2307/4117202
- Verdcourt B (1971a) Flora of Tropical East Africa: Annonaceae. 131 pp.
- Verdcourt B (1971b) Notes on East African Annonaceae. Kew Bulletin 25: 1–34. https://doi.org/10.2307/4103132
- Verdcourt B (1986) New Taxa of East African Annonaceae. Kew Bulletin 41(2): 287–297. https://doi.org/10.2307/4102932
- Verdcourt B (1996) Sanrafaelia, a new genus of Annonaceae from Tanzania. Garcia de Orta. Série Botanica 13: 43–44.
- Vollesen K (1980) Notes on Annonaceae from Tanzania. Botaniska Notiser 133: 53–62.
- Linné C von (1747) Flora Zeylanica. Holmia, 307 pp.
- Wang J, Chalermglin P, Saunders RMK (2009) The genus Dasymaschalon (Annonaceae) in Thailand. Systematic Botany 34(2): 252–265. https://doi.org/10.1600/036364409788606271
- Wang J, Thomas DC, Su YCF, Meinke S, Chatrou LW, Saunders RMK (2012) A plastid DNA phylogeny of Dasymaschalon (Annonaceae) and allied genera: Evidence for generic non-monophyly and the parallel evolutionary loss of inner petals. Taxon 61(3): 545–558. https://doi.org/10.1002/tax.613005
- Wang L-G, Lam TT-Y, Xu S, Dai Z, Zhou L, Feng T, Guo P, Dunn CW, Jones BR, Bradley T, Zhu H, Guan Y, Jiang Y, Yu G (2020) Treeio: An R package for phylogenetic tree input and output with richly annotated and associated data. Molecular Biology and Evolution 37(2): 599–603. https://doi.org/10.1093/molbev/msz240
- Watts DP, Potts KB, Lwanga JS, Mitani JC (2012) Diet of chimpanzees (Pan troglodytes schweinfurthii) at Ngogo, Kibale National Park, Uganda, 1. diet composition and diversity. American Journal of Primatology 74(2): 114–129. https://doi.org/10.1002/ajp.21016
- Weberling F, Hoppe JR (1996) Comparative morphological evaluation of inflorescence characters in Annonaceae. In: Reproductive Morphology in Annonaceae. Biosystematics and Ecology 10: 29–53.
- White F (1979) The Guineo-Congolian Region and its relationships to other phytochoria. Bulletin du Jardin botanique national de Belgique / Bulletin van de National Plantentuin van België 49: 11–55. https://doi.org/10.2307/3667815
- White F (1983) The Vegetation of Africa: A Descriptive Memoir to Accompany the Unesco/AETFAT/UNSO Vegetation Map of Africa. Unesco, Paris, 356 pp.
- White F (1993) The AETFAT chorological classification of Africa: History, methods and applications. Bulletin du Jardin botanique national de Belgique / Bulletin van de National Plantentuin van België 62: 225–281. https://doi.org/10.2307/3668279
- Wickham H (2016) ggplot2: Elegant graphics for data analysis. Springer-Verlag, New York. https://doi.org/10.1007/978-3-319-24277-4
- Xue B, Guo X, Landis JB, Sun M, Tang CC, Soltis PS, Soltis DE, Saunders RMK (2020) Accelerated diversification correlated with functional traits shapes extant diversity of the early divergent angiosperm family Annonaceae. Molecular Phylogenetics and Evolution 142: e106659. https://doi.org/10.1016/j.ympev.2019.106659
- Yamagiwa J, Mwanza N, Yumoto T, Maruhashi T (1994) Seasonal change in the composition of the diet of eastern lowland gorillas. Primates 35(1): 1–14. https://doi.org/10.1007/BF02381481
- Yu G, Smith DK, Zhu H, Guan Y, Lam TT-Y (2017) ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods in Ecology and Evolution 8(1): 28–36. https://doi.org/10.1111/2041-210X.12628
- Zhang C, Rabiee M, Sayyari E, Mirarab S (2018) ASTRAL-III: Polynomial time species tree reconstruction from partially resolved gene trees. BMC Bioinformatics 19(S6): e153. https://doi.org/10.1186/s12859-018-2129-y
- Zizka A, Rydén O, Edler D, Klein J, Perrigo A, Silvestro D, Jagers SC, Lindberg SI, Antonelli A (2021) Bio-Dem, a tool to explore the relationship between biodiversity data availability and socio-political conditions in time and space. Journal of Biogeography 48(11): 2715–2726. https://doi.org/10.1111/jbi.14256
Supplementary materials
Phylogenetic tree of the Monodoreae inferred with ASTRAL, based on 334 nuclear genes trees
Data type: pdf
Explanation note: The branch support is given as pie charts of the quartet support (QS) such as the black proportion represents the percentage of quartets in the gene trees agreeing with the branch in the species tree. The light blue box encloses the Monodoreae tribe. The name of the specimens is composed of a unique identifier, followed by the species (or variety/subsepecies) name, followed by the 3 first letters of the name of the collector and the collection number. For the details on the specimens see Suppl. material
RAxML maximum likelihood tree of the Monodoreae, based on a concatenated supermatrix of 334 nuclear genes. Bootstrap support values (in %) are given at the nodes
Data type: pdf
Explanation note: The light blue box encloses the Monodoreae tribe. The name of the specimens is composed of a unique identifier, followed by the species (or variety/subsepecies) name, followed by the 3 first letters of the name of the collector and the collection number. The identification of the specimens follow the taxonomic changes treated in this study. For the details on the specimens see Suppl. material
Details of the vouchers used in this study.
Data type: ods
Details of the specimens examined in this study.
Data type: tsv