Research Article |
|
Corresponding author: Werner Greuter ( w.greuter@bgbm.org ) Academic editor: Patrick Herendeen
© 2023 Rosa Gloria Rankin Rodríguez, Pedro Alejandro González Gutiérrez, Werner Greuter.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Rankin Rodríguez RG, González Gutiérrez PA, Greuter W (2023) A note on the type of Harpalyce (Fabaceae, Brongniartieae), with description of two new local endemic species from Cuba. PhytoKeys 225: 83-97. https://doi.org/10.3897/phytokeys.225.99321
|
Abstract
Two new species of Harpalyce are described from Cuba, H. revoluta sp. nov. from a serpentine area in the northern part of E. Cuba and H. marianensis sp. nov. from calcareous areas in the southern part of E. Cuba. Both have relatively small flowers, with an up to 6 mm long standard, and 2–3 mm long wings. Harpalyce marianensis is further characterized by strongly suberous (corky) young branches of a spongy consistence, deeply furrowed longitudinally, and by leaflets covered with a particular type of orange, apparently disk-shaped, sessile glands abaxially. Harpalyce revoluta, moreover, has suborbicular or broadly elliptic leaflets with a strongly recurved or sometimes revolute margin, secondary veins inconspicuous on either side; the foliar glands, by their morphology and anatomy, are of a different type. An epitype is designated for the name Harpalyce and its type, H. formosa; the distribution of both new species and their close relatives is mapped, and an updated identification key is offered, to cater for all 16 currently recognised Cuban species.
Resumen
Se describen dos nuevas especies de Harpalyce para Cuba oriental, H. revoluta sp. nov. de las áreas de serpentina del norte y H. marianensis sp. nov. de áreas de formaciones calcáreas del sur. Ambas se caracterizan por tener flores pequeñas, con estandarte de hasta 6 mm de largo y alas de hasta 3 mm de largo. Harpalyce marianensis se distingue además por sus ramas jóvenes fuertemente suberosas, de consistencia esponjosa, profundamente surcadas longitudinalmente, y los folíolos cubiertos abaxialmente por un tipo particular de glándulas sésiles anaranjadas. Harpalyce revoluta, por su parte, tiene folíolos suborbiculares o anchamente ovales, con margen fuertemente recurvado o hasta revoluto, con venas secundarias inconspicuas en ambas caras, y glándulas foliares, por su morphología y anatomía, son de un tipo distinto. Se designa un epitipo para el nombre Harpalyce y su tipo, H. formosa. Se presenta un mapa de distribución de las nuevas especies y las relacionadas con ellas y se ofrece una clave de identificación actualizada, que incluye las 16 especies cubanas actualmente reconocidas.
Keywords
Brongniartieae, Eastern Cuba, Fabaceae, Harpalyce, leaf gland morphology, limestone substrate, serpentine
Palabras clave
Brongniartieae, Cuba oriental, Fabaceae, Harpalyce, morfología de glándulas foliares, serpentinas, substratos calcáreos
Introduction
Harpalyce DC. is one of those genera that, so to say, sprouts new species on a regular basis, with apparently increasing frequency. The phenomenon is not limited to Cuba, but can also be observed in the two other, disjunct areas in which the genus occurs: Central Mexico to Nicaragua in Central America, and large parts of Brazil (
In Cuba, from where 16 species (including the two present, new ones) are now known, Harpalyce was first collected by Charles Wright in the 1860s. The species described by
Materials and methods
The original and complementary literature dealing with Harpalyce (Fabaceae, Brongniartieae) was consulted. In addition to the ca. 170 materials studied earlier (
Leaf glands were studied using the stereo-zoom microscope system Olympus SZX16; the photographs were taken using the microscope’s camera Olympus DP72 and its CellSens Standard software. Cross cuts of the lamina were obtained by razor-cutting manually mature leaflets, previously soaked soap water at room temperature. Unstained cuts in aqueous solution were studied at 200× and 400× magnification by optical microscopy using a Carl Zeiss Axioscop instrument.
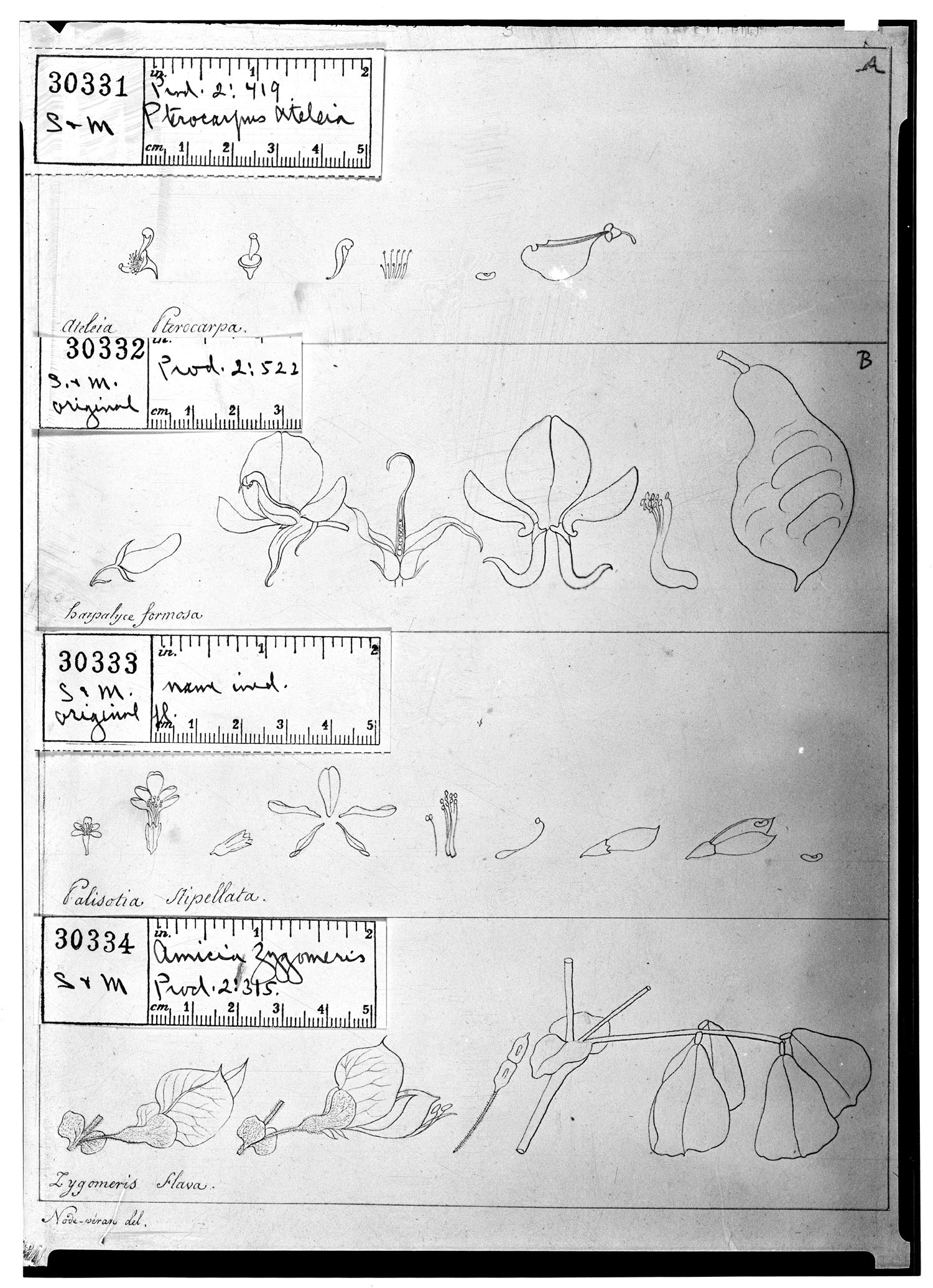
Notes on the type of and author citation for the name Harpalyce
While searching the literature for potentially relevant information, we came across
Harpalyce formosa DC., Prodr. 2: 523. 1825. Holotype [or, if lectotype, designated as such by
A further point of note clarified by
Description of two new Cuban species of Harpalyce
Harpalyce marianensis , sp. nov.
Type material
Holotype. Cuba – Guantánamo Prov. • San Antonio del Sur, Abra de Mariana en el barranco, 5 km al noroeste de San Antonio del Sur; [20.08828°N, 74.85846°W]; A. Álvarez de Zayas et al. HFC 43077; montes secos, caliza; 11 May 1980; JE 28983.
Isotypes. Cuba • Same collection data as for preceding; B 100364878 [http://herbarium.bgbm.org/object/B100364878]; HAJB 1289, 1290; JE 28984.
Other material examined
Cuba – Guantánamo Prov. • San Antonio del Sur, manigua costera cerca de Playa Baitiquirí; [20.02482°N, –74.85226°W]; May 1968, Bisse & Köhler HFC 9259; HAJB, JE; Baitiquirí, en el camino a la Mina del Yeso; [20.02482°N, 74.85226°W]; 11 Apr. 1972; Bisse & Berazaín HFC 21818; HAJB, JE; Abra de Mariana, loma al oeste del barranco; [20.08828°N, 74.85846°W]; 6 Feb. 1978; Bisse et al. HFC 36575; B 100362446, HAJB, JE; Abra de Mariana, loma al oeste del Abra; [20.08828°N, 74.85846°W]; 9 Feb. 1979; Berazaín et al. HFC 39129; B 100361112, HAJB, JE.
Shrubs or small trees; young branches very suberous (corky), with a spongy appearance, deeply longitudinally ridged (Fig.



Phenology
Collected in flower and with fruits in April and May.
Etymology
Named after the type locality, Abra de Mariana.
Distribution
Southern part of E Cuba, province Guantánamo, municipality San Antonio del Sur: Abra de Mariana and Baitiquirí. Grows in dry scrub vegetation on limestone substrate. Fig.

Affinities and diagnostic features
Harpalyce marianensis has morphological affinities with H. revoluta (see below) and other Cuban species that have a relatively small (≤ 8–9 mm in length or diameter), orbicular to broadly elliptic standard and free keel petals, such as: Harpalyce ekmanii Urb., H. villosa Britton & P. Wilson, and H. baracoensis Borhidi & O.Muñiz, particularly with H. ekmanii, with which it also shares glabrous or subglabrous leaflets; however, H. ekmanii has larger leaflets (2.5–7.5 × 1.5–3 cm vs 1.2–3.3 × 0.8–1.5 cm). Harpalyce marianensis has pronounced suberous young branches with a spongy consistence and deep longitudinal ridges, a feature that is not found in H. ekmanii. Also, H. marianensis grows in dry scrub on limestone in the southern part of E Cuba, whereas H. ekmanii grows in rainforest, pine woods or dry scrub on ultramaphic soils such as serpentines in the northern portion of E Cuba.
Sterile plants or specimens of Harpalyce marianensis might be confused with H. maisiana León & Alain, a species that also occurs in limestone areas of southern E. Cuba, but these two can be distinguished from each other through the colour of the glands on the adaxial face of the leaflets, yellow in H. maisiana vs orange in H. marianensis. When in flower, they can be easily told apart: the standard is oblanceolate, 1 × 0.2–0.3 cm and apically slightly emarginate in H. maisiana but orbicular or broadly elliptic, 5–6 mm in diameter or length and apically rounded in H. marianensis; the keel petals are partially connate in the apical third and 1.2–1.3 × 0.2–0.3 cm in H. maisiana, but totally free and 2.3–2.5 × 0.2–0.3 cm in H. marianensis.
Harpalyce revoluta , sp. nov.
Type material
Holotype. Cuba – Holguín Prov. • Cuchillas de Moa; alrededores del aserrío La Melba; A. Álvarez de Zayas et al. HFC 42194; charrascales altos con Bonnetia cubensis; [20.51944°N, 74.81778°W]; alt.450–500 m; 28 Apr. 1980; B 100364586 [http://herbarium.bgbm.org/object/B100364586].
Isotypes. Cuba • Same collection data as for preceding; HAJB 1286-1288, JE 28985.
Other material examined
Cuba – Holguín Prov. • Moa; La Melba; charrascal cerca del aserrío; [20.51944°N, 74.81778°W]; Mar. 1968; Bisse & Köhler HFC 7066; HAJB, JE; La Melba, charrascal cerca del aserrío; [20.51944°N, 74.81778°W]; Mar. 1968; Bisse & Köhler HFC 7626; HAJB, JE; La Melba; charrascal cerca del aserrío; [20.51944°N, 74.81778°W]; 22 Dec. 1968; Bisse & Lippold HFC 11062, 11329; HAJB, JE; Cayo Probado; orillas de las cabezadas del río Jiguaní; [20.48864°N, 74.82188°W]; 3 Apr. 1972; Bisse & Berazaín HFC 21968; B 100462873, HAJB, JE; alrededores del aserrío La Melba; [20.51944°N, 74.81778°W]; 25 Apr. 1981; Bisse et al. HFC 44956; B 100374694, HAJB, JE.
Shrubs or small trees; young branches pubescent, old branches (Fig.


{kind=link}
Phenology
When collected in late April, the type material was in flower and with immature fruits. Some other specimens seen, collected in April as well, similarly were in flower and with immature fruits.
Distribution
Northern part of E Cuba, province Holguín. Grows in rainforest, pine woods or xeromorphic scrub on ultramaphic substrate (serpentine). Fig.
Etymology
The epithet refers to the characteristic, revolute leaflet margin.
Affinities and diagnostic features
Harpalyce revoluta, shows morphological affinities with H. marianensis (see above) and other Cuban species that have a relatively small (≤ 8–9 mm in length or diameter), orbicular to broadly elliptic standard and free keel petals, particularly with H. ekmanii, with which it shares glabrous or subglabrous leaflets; however, in H. ekmanii the leaflets are much larger (2.5–7.5 × 1.5–3 cm vs 0.6–2 × 0.5–1 cm), with flat (vs recurved to revolute) margins, and the sessile glands on the abaxial leaf face are fairly dense, less spaced. The size and shape of the standard relates H. revoluta to H. villosa, but the latter has leaflets that, particularly when young, are densely hairy abaxially, not glabrous except on the loosely hairy midvein, as in H. revoluta.
An update of the identification key for Cuban Harpalyce
In their recent survey of Cuban Harpalyce, Rankin and González (2021) presented a key for identifying the species they recognise. To account for the present, additional species, that key needs some modification. That key is reproduced here, with its 8th dichotomy changed and expanded to account for the new species.
| 1 | Leaflets (especially when young) densely hairy, tomentose or pubescent abaxially, indumentum obscuring glands until leaflets are older and hairs are less dense | 2 |
| – | Leaflets glabrous, subglabrous or with scattered hairs, these never obscuring glands | 7 |
| 2 | Bracts leaf-like (foliose), 1–1.5 cm long | Harpalyce toaensis |
| – | Bracts small, linear or triangular, < 3 mm long | 3 |
| 3 | Leaflets (on dried specimens) green or greyish green adaxially, abaxially covered with a brownish yellow tomentum | 4 |
| – | Leaflets (on dried specimens) brown or dark grey adaxially, abaxially pubescent or puberulous, with brown or reddish brown hairs | 5 |
| 4 | Leaflet margin recurved or flat, or in some leaflets revolute; inflorescence 1–6-flowered | Harpalyce nipensis |
| – | Leaflet margin consistently revolute; inflorescence always 1-flowered | Harpalyce acunae |
| 5 | Petiolule of terminal leaflet 1–3 mm long | Harpalyce villosa |
| – | Petiolule of terminal leaflet 4–10 mm long | 6 |
| 6 | Leaflets 5–11 per leaf; secondary veins conspicuous abaxially | Harpalyce alainii |
| – | Leaflets 11–17 per leaf; secondary veins inconspicuous abaxially | Harpalyce baracoensis |
| 7 | Keel petals completely free from each other | 8 |
| – | Keel petals connate for most of their length or at least in apical third | 11 |
| 8 | Standard 1 cm in diameter; wings c. 1.5 cm long | Harpalyce cristalensis |
| – | Standard 0.35–0.9 cm in diameter or length; wings 0.2–0.7 cm long | 9 |
| 9 | Leaflets mostly suborbicular or ± broadly elliptic, their margin recurved to strongly revolute; secondary veins inconspicuous on both faces | Harpalyce revoluta |
| – | Leaflets elliptic to narrowly elliptic, oblong or oblong-elliptic, their margin flat or almost so; secondary veins commonly conspicuous, at least abaxially | 10 |
| 10 | Young branches very suberous, with a spongy consistence, deeply longitudinally ridged; leaflets 1.2–3.3 × 0.8–1.5 cm | Harpalyce marianensis |
| – | Young branches without a spongy appearance, not longitudinally ridged; leaflets 2.5–7.5 × 1.5–3 cm | Harpalyce ekmanii |
| 11 | Standard ≤ 1.5 cm long | 12 |
| – | Standard ≥ 2 cm long | 13 |
| 12 | Petioles 1.5–2 cm long; standard c.1.5 × 0.8–1 cm; wings c. 1.5 cm long, keel petals c. 1.7 cm long | Harpalyce borhidii |
| – | Petioles 0.6–1.2 cm long; standard c. 1 × 0.2–0.3 cm; wings 1–1.1 cm long, keel petals 1.2–1.3 cm long | Harpalyce maisiana |
| 13 | Leaflets subopposite or alternate; wings black toward base, keel petals mostly black, with a yellow margin; legume 4–5 cm long | Harpalyce greuteri |
| – | Leaflets opposite; wings and keel petals red, orange, yellow or white; legume either up to 2.3 cm or at least 5 cm long | 14 |
| 14 | Legume 5–6.5 cm long | Harpalyce macrocarpa |
| – | Legume 1–2.3 cm long | 15 |
| 15 | Leaflets abaxially with well-spaced glands, these not touching each other | Harpalyce cubensis |
| – | Leaflets abaxially with dense, partly contiguous or confluent glands | Harpalyce suberosa |
Discussion
As we stated in the introduction, the two new species here described are likely not the last addition to the genus. There are incomplete specimens in the Cuban herbaria, HAC and HAJB in particular, that in all likelihood represent new, undescribed species, which we shall duly name as soon as we succeed in relocating them in the field and collect complete, fertile material; furthermore, in view of the species definition we here adopt, following the tradition established by other authors, any newly discovered Harpalyce population is likely to represent a new taxon.
The question may be asked legitimately: is such a narrow species definition practical and defendable? We ignore whether our species are valid under a biological species concept. For all we know, they might all, or at least some of them, be freely interbreeding when brought together. Without much experimental work involving artificial crossing, it is impossible to know whether and to which extent evolutionary divergence has succeeded in establishing genetic barriers and cross-incompatibility between morphologically distinct populations. We are similarly ignorant of the extent of genetic isolation, if any, for many and indeed most species currently recognised in polymorphic Cuban genera. It is therefore legitimate to recognise morphologically distinguishable populations as separate taxa, and currently, no workable alternative to this course is available.
The role of small, isolated populations in evolution and speciation in higher plants has been studied by Runemark in the Aegean archipelago of Greece. He has coined the expression “reproductive drift” (
Concerning the general evolutionary context in which the Cuban populations are to be seen, it is useful to be mindful of the results and conclusions of recent phylogenetic studies (
A further promising field of additional studies that we propose to follow up in the near future has opened up through the preliminary study of leaf glands of the new species here described. We found that the slight but obvious difference of the gland pattern they presented was based on obvious structural differences, to be observed on leaf transects, which have led to the definition of two distinct gland types that we have named “type A” and “type B”. Screening the remaining species of the genus for the presence in them of these gland types, and of possible further types yet to be defined, is a promising field of study. Preliminary results of leaf surface studies indicate that the “type A” glands of Harpalyce marianensis are also found in H. ekmanii, H. maisiana as well as H. greuteri R.Rankin & P.A.González (not discussed here); whereas “type B” glands of H. revoluta occur in H. cristalensis Borhidi & O.Muñiz. The scattered glands of the leaves of H. villosa, H. baracoensis, and H. acunae Borhidi & O.Muñiz, hidden by dense indumentum, might represent yet another type.
Acknowledgements
We express our gratitude to the herbaria B and HAJB, their technicians and curators for facilitating the study of specimens; to Alelí Morales and Wildee Alonso from HAJB, who provided information on, and photos of, some specimens; to Bibiana Moncada, Berlin, for providing preparations and micrographs of the leaf surface and leaf transects; to Michael Rodewald who edited the original illustrations; to H. Manitz for the loan of Harpalyce specimens housed in JE; and to Eddy Martínez Quesada and Isis Hernández from Camagüey, Cuba, for generating the distribution map based on our data. The revision of materials in the herbaria B and JE was possible thanks to scholarships awarded by the Verein der Freunde des Botanischen Gartens und des Botanischen Museums Berlin-Dahlem to Pedro A. González in 2022. This contribution to the taxonomic revision of Harpalyce for “Flora de la República de Cuba” is one of the results of the Cuban national project PN223LH010-018 “Estudios taxonómicos y fitogeográficos en familias de la flora cubana”, which involves the National Botanical Garden of Cuba and other Cuban institutions. Thanks to the Stiftung Herbarium Greuter for bearing the publication cost.
References
- Arroyo MTK (1976) The systematics of the legume genus Harpalyce (Leguminosae: Lotoideae). Memoirs of the New York Botanical Garden 26: 1–80. https://openlibrary.org/books/OL11301894M/
- Borhidi A, Muñiz O (1977) Adiciones al conocimiento de la flora cubana. Ciencias Biológicas, Academia de Ciencias de Cuba 1: 131–146. https://openlibrary.org/books/OL11301894M
- Greuter W (2003) Introducción. In: Greuter W (Ed.) Flora de la República de Cuba. Serie A. Plantas vasculares. Fascículo 7. Gantner KG, Ruggell, iii pp.
- Grisebach A (1866) Catalogus florae cubensis, exhibens collectionem Wrightianam aliasque minores ex insula Cuba missas. Engelmann, Leipzig. https://doi.org/10.5962/bhl.title.177
- McVaugh R (2000) Botanical results of the Sessé & Mociño expedition (1787–1803). VII. A guide to relevant scientific names of plants. Hunt Institute for Botanical Documentation, Pittsburgh.
- Rankin Rodríguez R, González Gutiérrez PA (2021) Harpalyce greuteri (Leguminosae: Brongniartieae), a new species from eastern Cuba, with a synopsis of and key to the Cuban species of the genus. Willdenowia 51: 209–219. https://doi.org/10.3372/wi.51.51204
- Runemark H (1969) Reproductive drift, a neglected principle in reproductive biology. Botaniska Notiser 122: 90–129.
- Runemark H (1970) The role of small populations for the differentiation in plants. Taxon 19(2): 196–201. https://doi.org/10.2307/1217954
- São-Mateus WMB (2018) Filogenia molecular e tempo de divergência em Harpalyce (Leguminosae, Papilionoideae) e sinopse taxonômica da sect. Brasilianae. PhD tesis, Centro de Biociências, Universidade Federal do Rio Grande do Norte, Natal. https://repositorio.ufrn.br/handle/123456789/26522?mode=full
- São-Mateus WMB, Simon MG, de Queiroz LP, Jardim JG, Cardoso DBOS (2019) Two new species of Harpalyce (Leguminosae, Papilionoideae) from the Cerrado hotspot of biodiversity in Brazil. Kew Bulletin 74(4): e61. https://doi.org/10.1007/s12225-019-9845-y
- Standley PC (1922) Trees and shrubs of Mexico (Fagaceae-Fabaceae). Contributions from the US National Herbarium 23: 171–515. https://www.jstor.org/stable/i23491841
- Thiers B (2022+) Index herbariorum: a global directory of public herbaria and associated staff. New York Botanical Garden’s virtual herbarium. http://sweetgum.nybg.org/science/ih/ [Continuously updated, accessed Dec 2022]