





Citation: Appelhans MS, Wagner WL, Wood KR (2014) Melicope balgooyi Appelhans, W.L. Wagner & K.R. Wood, a new species and new record in Melicope section Melicope (Rutaceae) for the Austral Islands. PhytoKeys 39: 77–86. doi: 10.3897/phytokeys.39.7691
Melicope balgooyi, a new species of Melicope (Rutaceae) is described. It is known only from the Austral Islands in the South Pacific (French Polynesia). However, it is not closely related to the other two species previously known from the Austral Islands, which are part of Melicope section Vitiflorae. The new species belongs to Melicope section Melicope and is most closely related to species from New Zealand, the Kermadec Islands, and the Society Islands. The new species has alternate to sub-opposite leaves, which is a very rare arrangement in Melicope and has only been described for two other species of the genus so far.
Austral Islands, Bass Islands, Melicope, French Polynesia, Pacific biogeography, Rapa Iti, Rutaceae, IUCN Red List Category
In the course of phylogenetic and revisionary studies in Melicope J.R. Forst. & G. Forst., a new species was found, and along with its description, we discuss its biogeography and sectional placement. Melicope is the largest genus in Rutaceae, consisting of about 235 species divided into the four sections Lepta (Lour.) T.G. Hartley, Melicope, Pelea (A. Gray) Hook. f., and Vitiflorae T.G. Hartley (
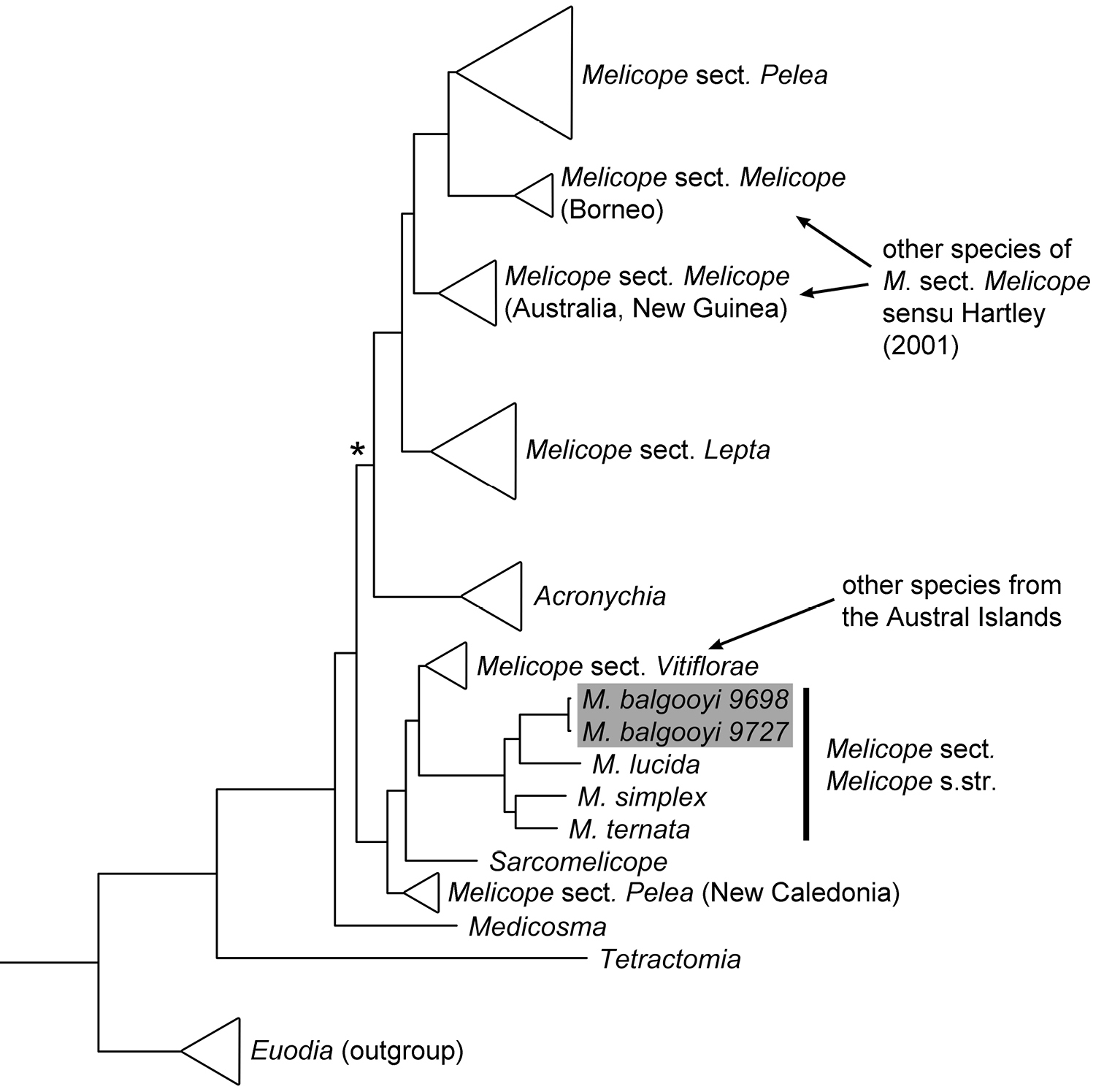
Phylogenetic placement of the newly described Melicope balgooyi Appelhans, W.L. Wagner & K.R. Wood is based on an analysis of combined chloroplast and nuclear data (redrawn from
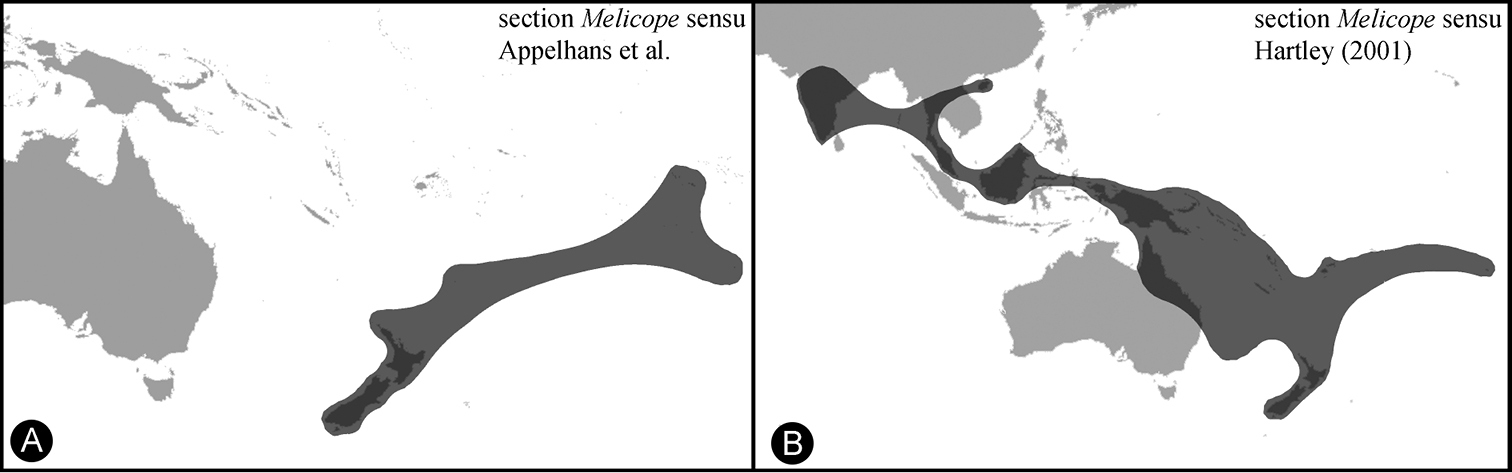
Distribution of Melicope section Melicope. A Melicope section Melicope sensu
urn:lsid:ipni.org:names:77140886-1
AUSTRAL ISLANDS: Rapa Iti, Maii, below rim near Pokumaru, 29 Apr 2002, K.R. Wood 9727 (holotype: PTBG-041326!, isotype: NY!).
This new species of Melicope differs from other species in that genus by the combination of alternate to sub-opposite leaves and oblanceolate leaves with a cordate base.
Shrub 50 to 150 cm of height; plants possibly dioecious; trichomes simple, greyish-white; branches brown-red and glabrous, 2–3 mm wide at third internode. Leaves with glandular dots, alternate to sub-opposite, unifoliolate, glossy dark green above with yellow or green-white midrib, 4.5–9 × 2.3–3.8 cm, petiole 0–1 mm long, terete, glabrous; blades sub-coriaceous, glabrous on both sides, obovate to oblanceolate, margin entire, apex rounded or obtuse, base cordate; venation brochidodromous, midrib prominulous or plane on both surfaces, secondary veins and veinlet reticulation prominulous, 10–17 secondary veins per side. Inflorescences unisexual, axillary, bracteate, several-flowered, up to 1.8 cm long, axes and bracts puberulent. Flowers unisexual, tetramerous; pedicel 1.3 to 2.2 mm long; sepals ovate to rounded, pellucid-dotted, glabrous or slightly ciliolate, 1.2 to 1.5 mm long, same size in staminate and pistillate flowers; petals ovate to elliptic, cream-green, pellucid-dotted, glabrous, 2.5 to 2.8 mm long, same size in staminate and pistillate flowers; stamens 8, glabrous; gynoecium 4-carpellate, glabrous, pellucid-dotted, 2 mm long, stigma peltate, young stigma white. Fruiting carpels and seeds not known.
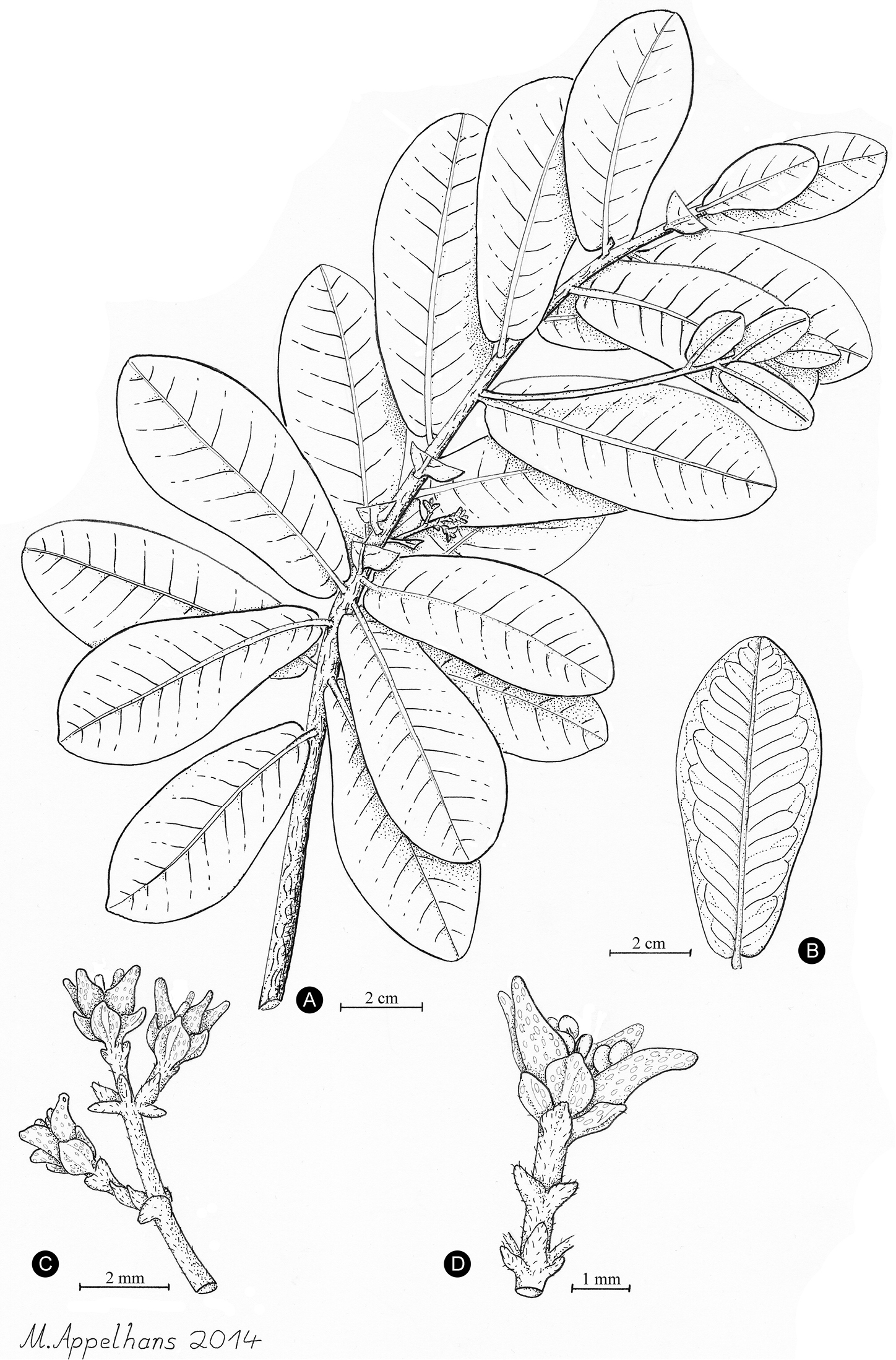
The newly described Melicope balgooyi Appelhans, W.L. Wagner & K.R. Wood. A Flowering branch B Leaf with detailed venation C Pistillate inflorescence [peduncle not shown in full length] D Single staminate flower. Based on K.R. Wood 9727 (PTBG, A–C) and K.R. Wood 9698 (PTBG, D).
(paratypes). Austral Islands. Rapa: Pokumaru summit region, upper windswept slopes of Maii, 21 Apr 2002, K.R. Wood 9698 (PTBG!, NY!).
Austral Islands (French Polynesia), only known from the type locality on slopes near the summit of Mount Pokumaru on Rapa; 550-580 m.
Melicope balgooyi was discovered around the tall spire-like peak of Pokumaru (Fig. 4b) between 550 and 580 m. The habitat is characterized by a windswept shrubland and forest that runs along and below an east to west ridgeline. The plant communities around Pokumaru are unique with small relictual patches of tropical montane cloud forest (TMCF) along with adjacent wet cliffs and steep slopes dominated by Freycinetia arborea Gaudich. Tree species in the TMCF zone which are associated with Melicope balgooyi include Fitchia rapense F. Br., Meryta choristantha Harms, Oparanthus coriaceus (F. Br.) Sherff, Carokia collenettei Riley, Metrosideros collina (J.R. Forst. & G. Forst.) A. Gray, Geniostoma rapense F. Br., and Weinmannia rapensis F. Br. Shrubs, vines, and herbs include Dianella intermedia Endl. var. punctata F. Br., Astelia rapensis Skottsb., Plantago rupicola Pilg., Alyxia stellata (J.R. Forst. & G. Forst.) Roem. & Schult., Freycinetia arborea, and Hebe rapensis (F. Br.) Garnock-Jones. Dominant ferns include Sphaeropteris medullaris (G. Forst.) Bernh., Alsophila stokesii (E.D. Br.) R.M.Tryon, Blechnum attenuatum (Sw.) Mett., Blechnum orientale L., Blechnum venosum Copel., Blechnum vulcanicum (Blume) Kuhn var. rapense E.D. Br., Polystichum rapense E.D. Br., Dicranopteris linearis (Burm. F.) Underw., Belvisia dura (Copel.) Copel., Doodia media R. Br., Elaphoglossum savaiense (Baker) Diels, and Davallia solida (G. Forst.) Sw. Less than 20 individuals of Melicope balgooyi are estimated to occur around this only known site.

A Habit of Melicope balgooyi Appelhans, W.L. Wagner & K.R. Wood in situ B Habitat of Melicope balgooyi around the steep precipitous slopes of Pokumaru, Rapa, Austral Islands.
The species is named in honor of Dr. Max M.J. van Balgooy, a specialist of the SE Asian flora. Dr. van Balgooy has annotated the herbarium sheets of this new species stating that it is an “unusual Rutaceae near Platydesma”. Phylogenetic studies (
The new species from Rapa differs from most Melicope species by its alternate to sub-opposite phyllotaxis. While most Melicope species are typically opposite-leaved, some species from all sections except Vitiflorae can have whorled leaves. This character state is most common on the Hawaiian Islands, where eight of the 52 species (incl. four species of Platydesma;
The placement of the new taxon in Melicope is assured by molecular phylogenetic data (
Two other species of Melicope [Melicope bracteata (Nadeaud) S.L. Welsh and Melicope margaretae (F. Br.) T.G. Hartley] have been described previously from the Austral Islands, but they are both members of Melicope section Vitiflorae (Fig. 1). Among other features, the new species differs most distinctly from Melicope bracteata and Melicope margaretae in stamen number (8) and oblanceolate leaves with a cordate base. Melicope bracteata and Melicope margaretae each have 4 stamens and usually elliptic leaves with a rounded or acute base.
The new taxon is connected to its closest relatives within Melicope section Melicope by its 8 stamens. Morphologically, and also phylogenetically, the new taxon mostly resembles the Tahitian Melicope lucida and Melicope tahitiensis. The often auriculate leaf base of many specimens of Melicope lucida and Melicope tahitiensis is similar to the cordate leaf base of the new taxon. The leaf shape among the three taxa is also similar; however, Melicope lucida and Melicope tahitiensis have acuminate to acute apexes as opposed to a rounded or slightly obtuse apex in the new taxon. Leaves of Melicope lucida and Melicope tahitiensis are petiolate, while those of the new taxon are sessile or subsessile. The new species further shares unisexual flowers and peltate stigmas with Melicope lucida and Melicope tahitiensis. The holotype specimen has pistillate flowers, while the paratype has staminate flowers, indicating the species may be dioecious like its closest relatives Melicope lucida and Melicope tahitiensis. However, it is important to note that the description of the new taxon is based on only two collections with a very low number of flowers, so that a definite statement about the sexual system of the species is not possible with the data at hand.
In order to differentiate between Melicope and several of its closely related genera on a morphological basis, fruit and seed characters are needed (
The Austral Islands are part of French Polynesia and are situated in the Southern Pacific. The Archipelago lies south of the Society Islands and consists of seven main islands of volcanic origin. Rapa is the second largest of these islands (i.e. 40 km2) and is about 5 million years old. The island is very rugged and is characterized by its steep central ridges, mist shrouded spires, and towering black basalt sea-cliffs. The highest peak is Mont Perau at about 650 m (
The affinities of Rapa’s flora are closely allied to New Zealand and Australia, yet with numerous exceptions. The high levels of endemic biological diversity in both the flora and fauna still puzzle many scientists because of the islands relatively small square area (
Three species of Melicope occur on the Austral Islands, which are the result of two independent colonization events. The newly described species is part of section Melicope and its closest relatives stem from Tahiti, the Kermadec Islands, and New Zealand. The two other species, Melicope bracteata and Melicope margaretae, are part of section Vitiflorae with an origin probably in the area of New Caledonia, Vanuatu, and Fiji based on the distributions of their closest relatives (
IUCN Red List Category. When evaluated using the World Conservation Union (IUCN) criteria for endangerment (
We would like to thank Timothy Flynn (PTBG) for providing leaf material for molecular work and a loan of specimens to GOET. Jean-Yves Meyer and Priscille Frogier of the Délégation à la Recherche, Polynésie Française, for their continued logistical support of research in French Polynesia. Timothy J. Motley (NYBG), organizer of the 2002 Rapa Expedition, and the National Geographic Society for partial funding. Steve Perlman (NTBG), Jean-Francois Butaud (Service du Développement Rural, Tahiti), Cerdan Faraire (Rapa), and Jean-Yves Meyer for field assistance. The first author would like to thank Anita Walsmit Sachs-Jansen (L) and Alice Tangerini (US) for sharing knowledge on scientific illustration. We thank María Elena Reiner-Drehwald (GOET) for scanning the line drawing. We appreciate careful editorial review by Nancy Khan (US) and we thank H.-J. Esser (M) for the helpful and constructive review of the manuscript.