





Citation: Souza IM, Funch LS, de Queiroz LP (2014) Morphological analyses suggest a new taxonomic circumscription for Hymenaea courbaril L. (Leguminosae, Caesalpinioideae). PhytoKeys 38: 101–118. doi: 10.3897/phytokeys.38.7408
Hymenaea is a genus of the Resin-producing Clade of the tribe Detarieae (Leguminosae: Caesalpinioideae) with 14 species. Hymenaea courbaril is the most widespread species of the genus, ranging from southern Mexico to southeastern Brazil. As currently circumscribed, Hymenaea courbaril is a polytypic species with six varieties: var. altissima, var. courbaril, var. longifolia, var. stilbocarpa, var. subsessilis, and var. villosa. These varieties are distinguishable mostly by traits related to leaflet shape and indumentation, and calyx indumentation. We carried out morphometric analyses of 14 quantitative (continuous) leaf characters in order to assess the taxonomy of Hymenaea courbaril under the Unified Species Concept framework. Cluster analysis used the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) based on Bray-Curtis dissimilarity matrices. Principal Component Analyses (PCA) were carried out based on the same morphometric matrix. Two sets of Analyses of Similarity and Non Parametric Multivariate Analysis of Variance were carried out to evaluate statistical support (1) for the major groups recovered using UPGMA and PCA, and (2) for the varieties. All analyses recovered three major groups coincident with (1) var. altissima, (2) var. longifolia, and (3) all other varieties. These results, together with geographical and habitat information, were taken as evidence of three separate metapopulation lineages recognized here as three distinct species. Nomenclatural adjustments, including reclassifying formerly misapplied types, are proposed.
Fabaceae, Detarieae, integrative taxonomy, nomenclatural changes
Hymenaea L. is a genus of caesalpinioid legumes with 14 species (
The current taxonomy of the genus is largely based on Lee and Langenheim’s (1975) revision. In addition to recognizing the 14 currently accepted species, they reduced several species to varietal rank. These polytypic species were viewed by
Hymenaea courbaril is the most widely distributed species of the genus, almost matching the geographic range of Hymenaea in the New World. It also has the greatest economic importance in the genus, due to the high quality of its wood and its resin, the latter being used by native populations as incense, cement, in the manufacture of varnishes, and for medicinal purposes. Its nutritive fruits are sought after by mammals and birds (
The species and varietal limits of Hymenaea courbaril, Hymenaea stigonocarpa, and Hymenaea martiana were investigated by
Delineating precise species boundaries is a key task in plant taxonomy. This process has direct impacts on society, as there is a growing demand for credible taxonomic information that allows us to conserve, manage, and understand natural biodiversity (
Analyses of morphometric data can be useful in objectively demonstrating species limits, especially when combined with molecular markers (
The problem of defining species limits in the polytypic Hymenaea courbaril complex is revisited here under the USC conceptual framework by exploring morphometric, geographical, and ecological patterns as lines of evidence for the existence of separate metapopulation lineages. Specifically, we sought to test Lee and Langenheim’s (1975) hypothesis that Hymenaea courbaril should be treated as a polytypic species with six varieties.
A total of 96 specimens of the Hymenaea courbaril complex were examined in this study (vouchers listed in Appendix 1). All analyzed materials were sheets from the following herbaria: B, CEN, CEPEC, HRB, HUEFS, IBGE, IPA, LAGU, M, MBM, NY, RB, SP, SPF, U, UB, and UC. We selected specimens that displayed branch ends with fully-developed (mature) leaves. This criterion avoided considering young leaves from the tips of the branches or leaves at the bases of the branches that are often much larger. Each specimen analyzed was considered an individual, and identifications strictly followed
The leaves in the group studied here are bifoliolate; the leaflets range from oblong to ovate or obovate, with rounded, acute or obtuse apices; the main vein is displaced towards the inner margin resulting in an asymmetrical base, the outer portion being wider than the inner and extending beyond the attachment to the petiolule (Figure 1). Fourteen quantitative (continuous) characters were examined (Table 1; Figure 1). Only leaf traits were measured and quantified, as most herbarium sheets lacked flowers and/or fruits. In any case, flower morphology is much conserved in the species studied and the herbarium material examined usually contained only incomplete or damaged flowers. Measurements were taken of two fully developed leaves per dried herbarium sheet, using a graduated ruler (precision 1 mm).
List of the quantitative leaf characters in specimens of the Hymenaea courbaril complex. Letters in the second column refer to measurements depicted in Figure 14.
| Leaf characters (cm) | Measurements in Figure 1 |
|---|---|
| 1 - Total length of the leaflet | A |
| 2 - Leaflet length /width ratio | A / B |
| 3 - Length of the distal sixth | E |
| 4 - Inner width in distal third | C |
| 5 - Outer width in distal third | D |
| 6 - Inner width in middle third | F |
| 7 - Outer width in middle third | G |
| 8 - Inner width in basal third | H |
| 9 - Outer width in basal third | I |
| 10 - Main vein displacement | (G–F) / (I–H) |
| 11 - Base extension (length of the base from the inner attachment of the petiolule) | J |
| 12 - Petiole length | M |
| 13 - Petiolule length | L |
| 14 - Distance between inner and outer attachment of the petiolule | K |
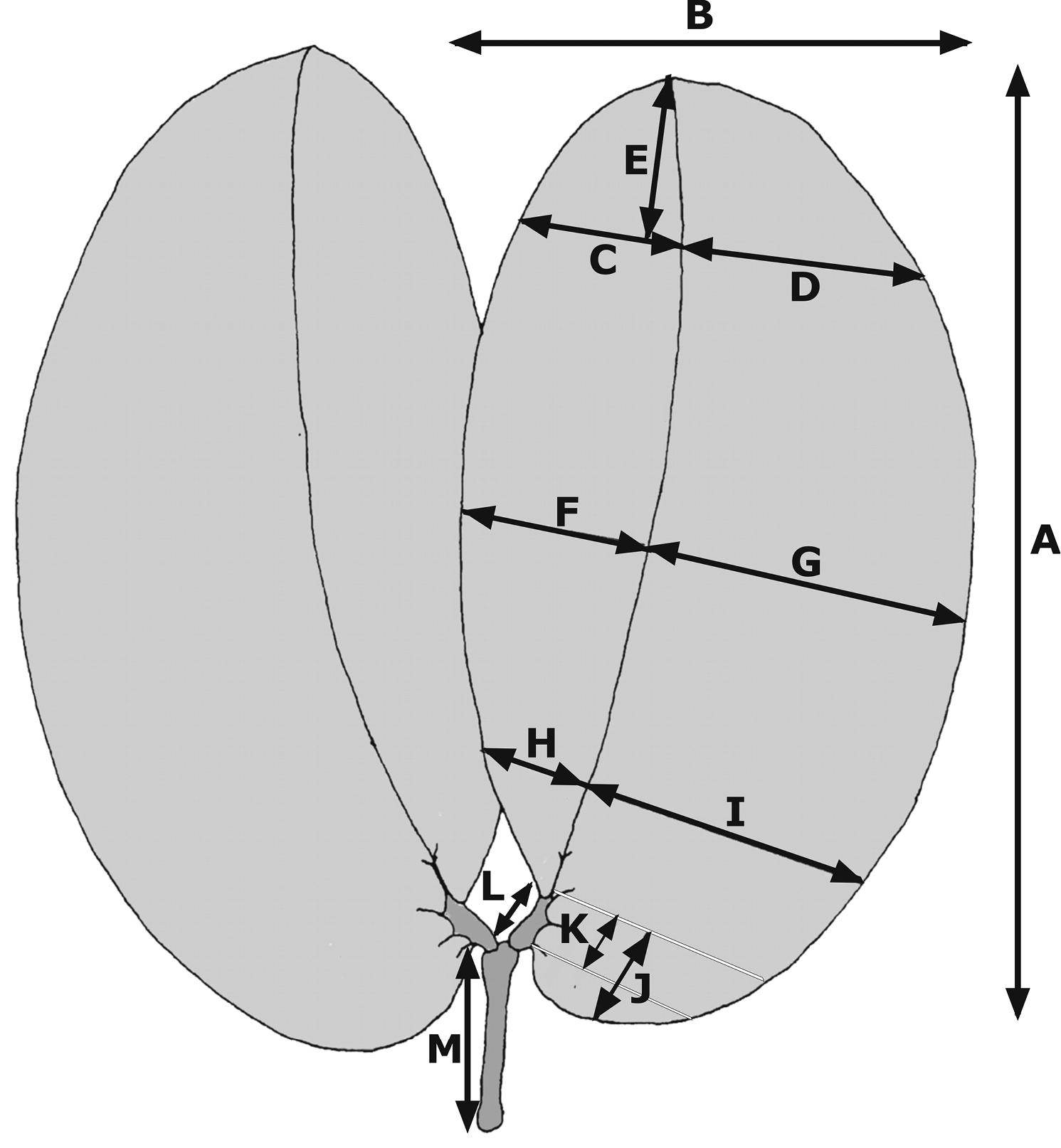
Quantitative leaf characters of specimens of the Hymenaea courbaril complex. Letters refer to the measurements described in Table 1.
All multivariate analyses were carried out using Past software (
Distribution maps of the specimens studied were prepared using DIVA-GIS software (Hijmanns et al. 2005), based on the geographic coordinates recorded on the herbarium sheet labels. For material lacking original coordinates, a central coordinate for the municipality was used as provided by the Species Link website (available at http://specieslink.org.br ).
Species limits were tested following the USC framework (
Both UPGMA and PCA recovered three major groups (Figure 2): Group 1 included all specimens of var. altissima; Group 2 all specimens of varieties courbaril, stilbocarpa, subsessilis, and the single specimen of villosa; and Group 3 all specimens of var. longifolia. Within Group 2, individuals of the different varieties did not cluster together and appeared intermixed in UPGMA, or formed highly overlapping groups in two first axes of PCA. The first PCA axis accumulated 88.3% of the total variance, with the two first axes summing 93% of the observed variation. Leaflet length was the trait that explained most of the variation found in first axis, and the three major groups were sorted mostly by leaflet size. These results indicate that Group 3 includes specimens with largest leaflets, and Group 1 the smallest leaflets (Figure 3).
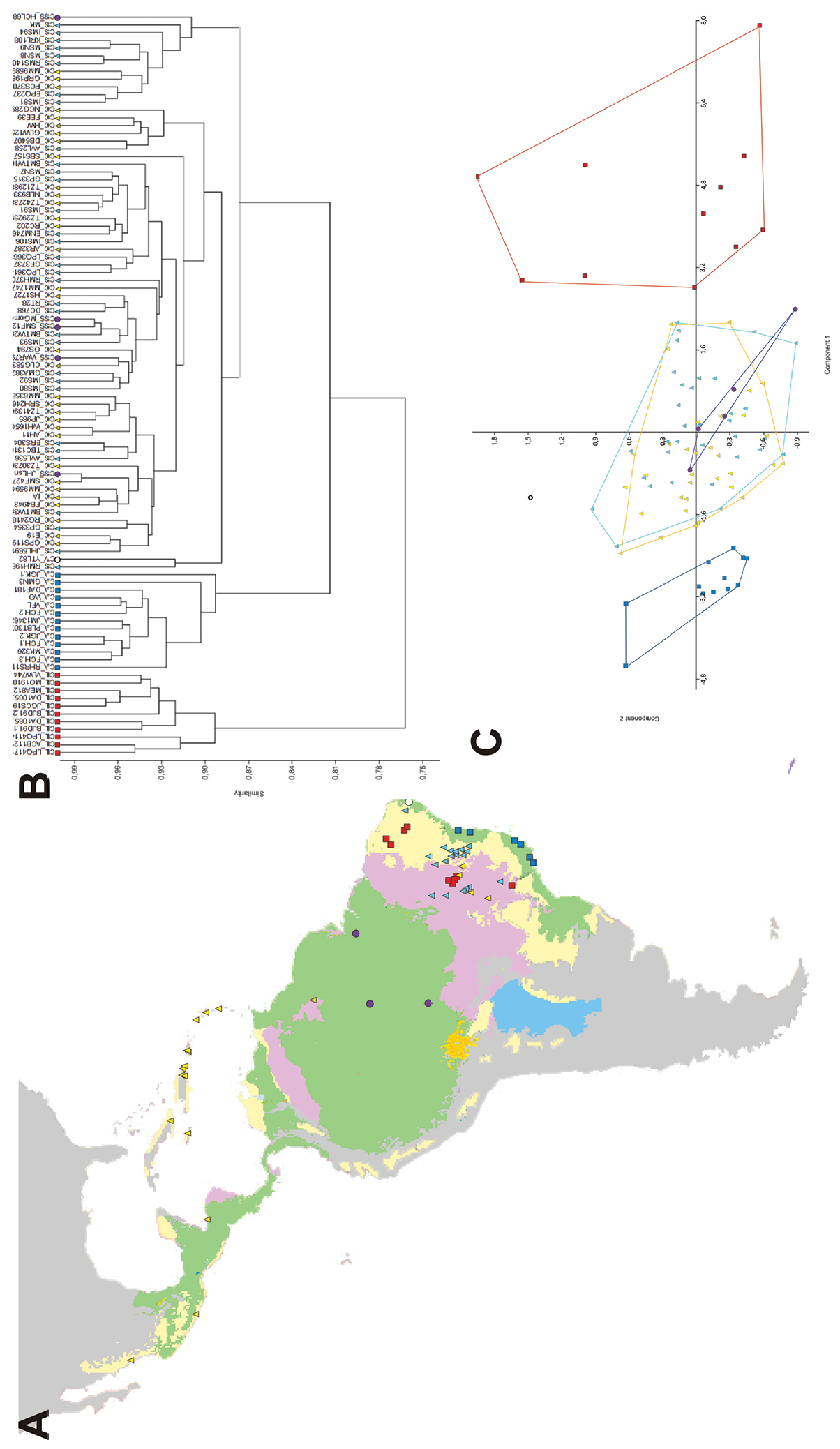
A Distribution of the Hymenaea courbaril complex in the Neotropics. The major biomes where this complex occurs are shown in color: wet forests (green), seasonally dry forests and woodlands (yellow), and savannas (purple). The varieties of Hymenaea courbaril recognized by
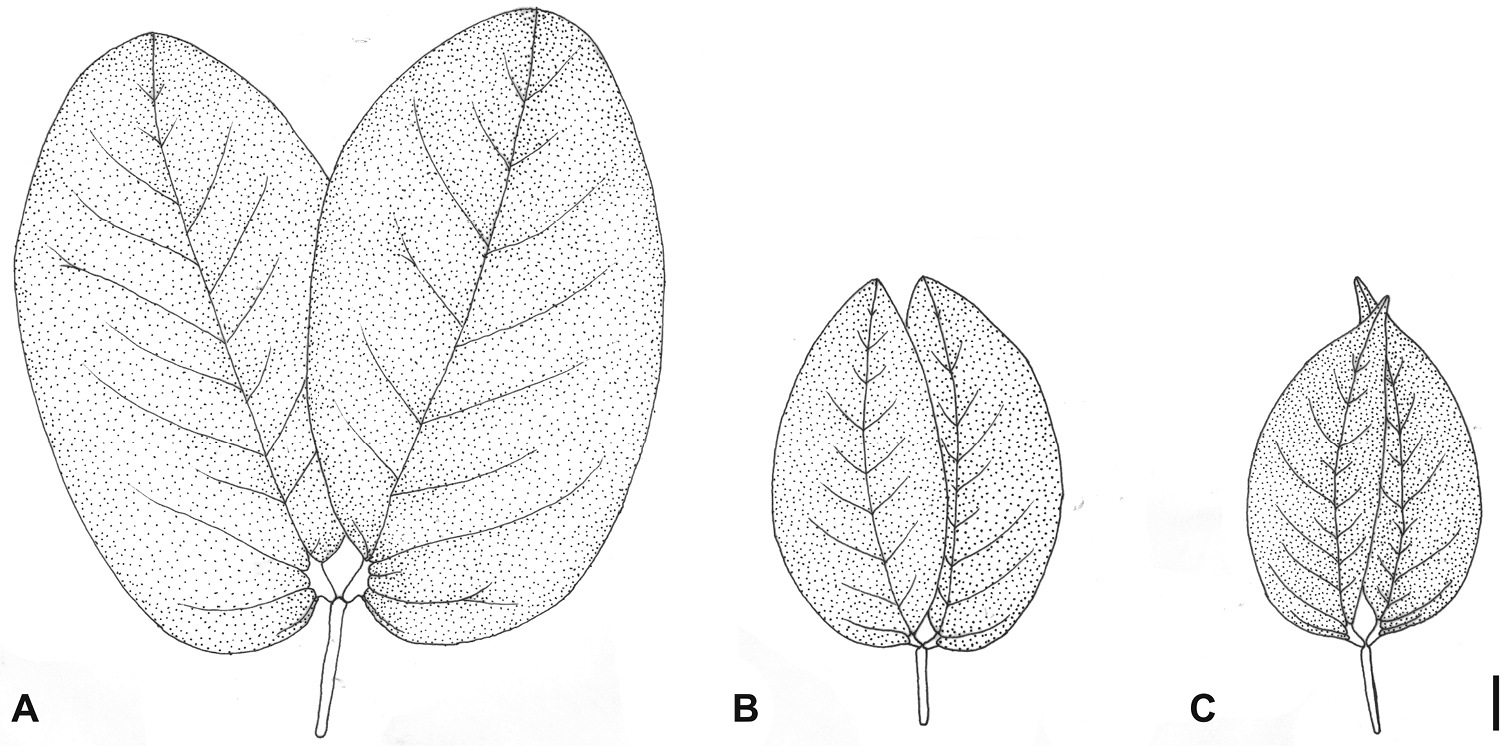
Leaf morphology of the species recognized in the Hymenaea courbaril complex: A Hymenaea longifolia B Hymenaea courbaril C Hymenaea altissima. Scale bar = 1 cm.
The results of the UPGMA and PCA were consistent with both ANOSIM and NPMANOVA. Comparisons of the varieties of Hymenaea courbaril showed significant differences (p < 0.05) between the varieties altissima and longifolia and all other varieties, but no significant differences between the varieties courbaril, silbocarpa, and subsessilis that clustered in Group 2 (Table 2). The same analyses comparing the three groups recovered in UPGMA and PCA found significant morphological differences between them (Table 3).
Results of ANOSIM and NPMANOVA testing the consistence of the varieties of Hymenaea courbaril, using 10, 000 permutations and Bray-Curtis distances. ANOSIM R-values are given above the diagonal and NPMANOVA F-values are given below the diagonal. Numbers between brackets are Bonferroni-corrected p-values. Significant differences are indicated by shadowed cells.
| stilbocarpa | courbaril | subsessilis | altissima | longifolia | |
|---|---|---|---|---|---|
| stilbocarpa | 0.09933 [0.0195] | -0.1343 [1] | 0.9325 [0.0015] | 0.8858 [0.0015] | |
| courbaril | 5.313 [0.0855] | 0.09375 [1] | 0.8524 [0.0015] | 0.9471 [0.0015] | |
| subsessilis | 0.8397 [1] | 3.296 [0.489] | 0.9972 [0.0045] | 0.9329 [0.0075] | |
| altissima | 79.79 [0.0015] | 57.97 [0.0015] | 49.86 [0.003] | 1 [0.0015] | |
| longifolia | 67.13 [0.0015] | 94.08 [0.0015] | 30.27 [0.006] | 235.5 [0.0015] |
ANOSIM and NPMANOVA (10, 000 permutations and Bray-Curtis distances) for the groups recovered in UPGMA and PCA analyses of 78 individuals of Hymenaea courbaril complex. Group 1 included all specimens of var. altissima, Group 2 all specimens of the varieties courbaril, stilbocarpa, subsessilis, and villosa, and Group 3 all specimens of var. longifolia. Values before the slash resulted from the analyses including var. villosa and after one excluding it. ANOSIM R-values are given above the diagonal and NPMANOVA F-values are given below the diagonal. Numbers in brackets are Bonferroni-corrected p-values. Significant differences are indicated by shadowed cells.
| Group 1 | Group 2 | Group 3 | |
| Group 1 | 0.8762 / 0.8771[0.0003 / 0.0003] | 1 / 1[0.0003 / 0.0003] | |
| Group 2 | 73.65 / 74.35[0.0003 / 0.0003] | 0.9045 / 0.9055[0.0003 / 0.0003] | |
| Group 3 | 235.5 / 235.5[0.0003 / 0.0003] | 84.47 / 85.58[0.0003 / 0.0003] |
The complex morphological variations found in Group 2 appear to reflect its wide geographic range and large genetic variability (
Leaflet indument distinguishes var. villosa from the remaining varieties of Group 2 (Lee and Andrade-Lima 1974, 1975). The ovary is also described as slightly pilose on one side, a condition not observed in other varieties of Hymenaea courbaril. The exclusion of var. villosa from the similarity analyses did not alter the results obtained with its inclusion (Table 3). The var. villosa is known from only two specimens from the eastern coastal areas of the states of Paraíba and Pernambuco (northeastern Brazil) where the range of var. stilbocarpa overlaps with that of Hymenaea martiana. This species (Hymenaea martiana) has tomentose leaflets and an ovary with a tuft of trichomes near its base. The rarity of Hymenaea courbaril var. villosa, as well as the transitional nature of the leaflets and ovary indumentation in relation to Hymenaea martiana and the other varieties of the Hymenaea courbaril complex clustered in Group 2, all suggest that var. villosa could represent a hybrid between Hymenaea martiana and Hymenaea courbaril.
The habitats of the taxa of Group 2 include mostly tropical ombrophilous alluvial (gallery) forests in areas subject to seasonally dry climates from Mexico and the Caribbean islands to central Brazil, but not the Amazonian region (var. courbaril and stilbocarpa), tropical ombrophilous lowland forests in central and eastern Amazon basin (var. subsessilis), or tropical ombrophilous submontane forests in the northeastern Brazilian state of Paraíba (var. villosa) (Figure 2).
The var. altissima (Group 1) constitutes a morphologically, geographically and ecologically consistent taxon. It is distinguished from the other varieties of Hymenaea courbaril complex by having smaller, falcate and acuminate leaflets, not exceeding 6.5 cm long on the flowering branches (Table 4). The leaflets of this taxon have a thinner texture than the remaining varieties, allowing its tertiary venation to appear as raised and reticulate veins. The flowers of var. altissima are also the smallest within the Hymenaea courbaril complex, measuring less than 15 mm long (
Morphological comparison between Hymenaea courbaril, Hymenaea altissima, and Hymenaea longifolia.
| Hymenaea courbaril | Hymenaea altissima | Hymenaea longifolia | |
|---|---|---|---|
| Leaflet outline | Elliptic or ovate, not or slightly falcate | Falcately elliptic | Oblong or narrowly elliptic, not falcate |
| Inner margin of the leaflet | Straight or slightly concave | Strongly concave | Straight |
| Outer base of the leaflet | Slightly rounded | Truncate | Rounded |
| Apex | Acute, rarely obtuse | Abruptly acuminate | Obtuse, rarely rounded |
| Petiole length (cm) | 1.1–1.7 (– 2) | 1–1.3 (– 1.7) | 2.6–3 |
| Petiolule length (mm) | 4–5 | 3–4 (– 5) | 7–8 |
| Leaflet size (cm) | 6–9 (– 10.2) × 2.8–4 (– 5.1) | 4–5.7 (– 6.4) × 2.0–2.5 | 10.1–12.5 (–15.4) × 4.5–6.5 (– 6.9) |
| Flower length (cm) | 2.2–3.1 | 2–2.7 | 2.6–3.8 |
| Fruit shape | Cylindrical, rounded in cross section | Cylindrical, rounded in cross section | Oblongoid, compressed in cross section |
| Seeds per fruit | Mostly > 4 | Mostly 1–2 | Mostly > 4 |
Var. longifolia (Group 3) comprises the morphs with largest leaflets. This variety also differs from the remaining groups by having oblong leaflets with an obtuse apex, clawed petals, and fruits compressed and enlarged toward the apex (Table 4). It occupies a very particular habitat in submontane tropical drought-deciduous thorny forests in the Caatinga and Cerrado phytogeographical domains that extend from western Bahia and Piauí states to the Araripe mountain range in the border area between the states of Ceará and Pernambuco (Figure 2). It was first proposed as a variety of Hymenaea splendida Vogel.
In addition to morphological distinctions, ecological and geographical information can be used for interpreting the three groups recovered by UPGMA and PCA as separately evolving lineages that would be considered different species under the USC framework (see Taxonomy section). With respect to the variation in Group 2, studies of wider samplings, including especially the collections made after Lee and Langenheim’s (1975) work, shows that the diagnostic features used for defining the varieties included in Group 2 have more complex variations. Indumented leaflets, for example, the diagnostic character for var. villosa, can be found to varying degrees in var. stilbocarpa. A short stipe was used as a diagnostic feature of var. subsessilis, but this trait is variable and grades into var. courbaril. Finally, the distinctions between vars. courbaril and stilbocarpa rely on the types and colors of the calyx lobe indumentation, traits that tend to change during the duration of the flower. Thus, a more parsimonious way to treat the taxonomy of Group 2 is to consider it a variable species, without recognizing varieties.
Large trees, to 30 m tall. Petiole 1.1–1.7 (–2) cm long; petiolule 4–5 mm long; leaflets 6–9 (–10.2) × 2.8–4 (–5.1) cm, elliptic, rarely ovate, straight or slightly falcate due to central vein with an angle c. 10°, apex acute or obtuse, rarely acuminate, base acute along the inner margin and rounded along the outer margin, distance from the inner margin to central vein 7–10 mm in the basal region. Flower buds 2.1–2.5 cm long; flower 2.2–3.1 cm long; hypanthium 7–15 mm long; petals 1.1–2.0 cm long. Fruit cylindrical, of uniform width, apex mostly rounded and apiculate.
Hymenaea courbaril is defined here more narrowly than the circumscription adopted by
Nomenclatural problems with several names associated with Hymenaea courbaril were not amended by
Hymenaea confertifolia Hayne was based on material collected by Sellow and Olfers in Brazil (“Wächst in Brasilien (Olfers u. Sellow)”; Hayne 1830: table 9). Lee and Langenheim (1975: 88) stated that this name was based on Sellow 1025 and that the “holotype” in the B herbarium was destroyed. However, as Hayne (1830) did not refer to one particular specimen, all materials collected by Sellow or Olfers that can be linked with Hymenaea confertifolia should be considered syntypes. No such specimens can be found in the B herbarium, and were probably destroyed. A duplicate from B collected by Sellow was found in W annotated as Hymenaea confertifolia, and is designated here as the lectotype of this name.
Hymenaea retusa Willd. ex Hayne was published as a homotypic synonym of Hymenaea candolleana (Hayne 1830). It is thus illegitimate under the Article 52 of the International Code of Botanical Nomenclature (
Hymenaea courbaril var. obtusifolia Ducke was published based on a tree cultivated in the Pará Botanical Garden (now Museu Paraense Emílio Goeldi, Belém, state of Pará, Brazil).
Ducke (1925: 265) did not cite any specimen when describing Hymenaea courbaril var. subsessilis Ducke.
Large trees, up to 38 m tall. Petiole 1–1.3 (–1.7) cm long; petiolule 3–4 (–5) mm long; leaflets 4–5.7 (–6.4) × 2–2.5 cm, oval, narrow-elliptic or elliptic, strongly falcate, due to central vein with an angle c. 35°, apex acuminate, rarely acute, base acute along the inner margin and truncate along the outer margin, distance from the inner margin to central vein 4–5 mm in the basal region. Flower buds 1.5–2.5 cm long; flower 2–2.7 cm long; hypanthium 7–12 mm long; petals 1.2–1.6 cm long. Fruit cylindrical, of uniform width, apex mostly rounded and apiculate.
Hymenaea altissima shows the smallest leaflets and flowers in the species group related to Hymenaea courbaril (Table 4). Additionally, its leaflets are strongly falcate due to the strongly curved and displaced midvein, with an abruptly acuminate apex. The base of the leaflet is acute along the inner margin and truncate along the outer margin (Figure 3). The fruits are mostly cylindrical, as in Hymenaea courbaril, but usually shorter than those of this species (4–5 vs. 8–20 cm long) because they have only one or two seeds, while in Hymenaea courbaril they usually have six or more seeds. This species is restricted to the coastal rain forests of eastern Brazil, from southern Bahia State to São Paulo and Paraná states.
This species was described by
Lee and Langenheim (1975: 84) stated that the lectotype of Hymenaea altissima was designated by
urn:lsid:ipni.org:names:77140226-1
Medium-sized trees c. 5–12 (–18) m tall. Petiole 2.6–3 cm long; petiolule 7–8 mm long; leaflets 10.1–12.5 (–15.4) × 4.5–6.5 (–6.9) cm, elliptic, narrow-elliptic, or oblong, not falcate, due to central vein with an angle c. 24°, apex obtuse or acute, base acute or rounded along the inner margin and rounded or slightly rounded along the outer margin, distance from the inner margin to central vein 10–11 mm in the basal region. Flower buds 2.2–3.3 cm long; flower 2.6–3.8 cm long; hypanthium 9–14 mm long; petals 1.3–2.1 cm long. Fruit oblongoid, compressed, wider at distal region, apex slightly truncate and apiculate.
Hymenaea longifolia, as circumscribed here, is characterized by large leaflets, both longer and wider than those of the related species Hymenaea courbaril and Hymenaea altissima, with longer petioles and petiolules (Table 4). The leaflets are elliptic or oblong with obtuse (rarely acute) apices and rounded bases along the outer margin (Figure 3). This species occurs in Seasonally Dry Forests within the Caatinga and Cerrado phytogeographical domains in northeastern Brazil, from Ceará State to the northern portion of Bahia State.
It was first described by
This paper was part of IMS’s Master Dissertation at the Graduate Program in Botany of the Universidade Estadual de Feira de Santana. We are grateful to the curators of the cited herbaria for loans of specimens or for making available digital images of their collections, to Roy R. Funch for the English revision, and to the two anonymous reviewers for their contributions to the text. IMS was supported by a CAPES grant and LPQ by a CNPq-Pq1C grant. This work was sponsored by Programa de Pesquisa em Biodiversidade do Semi-árido (PPBIO), Sistema Nacional de Pesquisa em Biodiversidade (SISBIOTA, processes CNPq 563084/2010-3 and FAPESB PES0053/2011), and the Programa de Capacitação em Taxonomia (PROTAX, CNPq process 562354/2010-7). IMS was supported by a CAPES grant and LPQ by a CNPq Pq 1C grant.
Materials examined for the morphometric study of the Hymenaea courbaril complex. Nomenclature follows Lee and Langenheim (1975).
Hymenaea courbaril var. altissima–BRAZIL, Bahia, Ilhéus, R.H.R.Sambuichi 1169 (CEPEC). Esprírito Santo, Linhares, D.A.Folli 181 (RB). Rio de Janeiro, Nova Iguaçu, W.Dias s/n (HUEFS); Resende, J.G.Kuhlmann 179 (RB); Rio de Janeiro, J.G.Kuhlmann s/n (RB). São Paulo, Moji das Cruzes, P.L.B.Tomasulo 307 (SP); São Paulo, F.C.Hoehne s/n (SP, SPF); J.Mattos 13463 (SP); M. Kirizawa 326 (SP); V.F.Lima s/n (SP).
Hymenaea courbaril var. courbaril–BRAZIL, Bahia, Baianópolis, S.B. da Silva 157 (HRB). Bom Jesus da Lapa, S.M. de Faria 427 (RB). Goiás, Niquelândia, G.P.Silva 1192 (CEN); Santa Bárbara de Goiás, A.Raw 3287 (UB). COLOMBIA, Santa Maria, Espina 19 (UC). CUBA, Santiago de Cuba, Mella, I.Arias s/n (B). EL SALVADOR, s/l, R.Cruz 202 (LAGU); P.C.Standley 3705 (UC). HONDURAS, Morazan, Jicarito, G.L.Webster 12529 (F). MEXICO, Acapulco, W. Humboldt s/n (B). PANAMA, Barro Colorado, N.C.Garwood 2891A (F); O.Shattuck 794 (UC). SURINAME, Boschreserve, F.Bureau 4943 (U).
Hymenaea courbaril var. longifolia–BRAZIL, Bahia, Barreiras, L.P. de Queiroz 4114 (HUEFS); Formosa do Rio Preto, D.Alvarenga 1065 (RB); B.J.Dias 91 (IBGE, SP); L.P. de Queiroz 4171 (HUEFS). Ceará, Brejo Santo, J.G.Carvalho-Sobrinho 1925 (HUEFS); Missão Velha, Academia Brasileira de Ciências 1121 (IPA). Piauí, Brasileira, M.E.Alencar 812 (HUEFS); Campo Maior, M.Oliveira 1910 (IPA). São Paulo, Bauru, V. de L. Weiser 744 (RB).
Hymenaea courbaril var. stilbocarpa–BRAZIL, Bahia, Caetité, L.P. de Queiroz 3614 (HUEFS); Gentio do Ouro, K.R.B.Leite 108 (HUEFS); E.R. de Souza 304 (HUEFS); R.Tourinho 28 (HUEFS); Jaguaripe, E.N. de Matos 746 (HUEFS); Livramento do Brumado, R.M.Harley 19889 (IPA); L.P. de Queiroz 3663 (HUEFS); Miguel Calmon, E.P.Queiroz 2372 (MBM); Mucugê, R.M.Harley 3702 (HUEFS); Palmeiras, M. de S. Nunes 7, 8, 9 (HUEFS); I.M.Souza 80, 81, 91, 92, 93, 94, 106 (HUEFS); Rui Barbosa, D.Cardoso 768 (HUEFS); Vitória da Conquista, J.H.Langenheim 5641 (IPA). Goiás, Alto Paraíso de Goiás, T.B.C. 1310 (HUEFS); Colinas do Sul, B.M.T.Walter 1038 (HUEFS); Minaçu, B.M.T.Walter 3570 (HUEFS); Niquelândia, B.M.T.Walter 2515 (HUEFS). Minas Gerais, Araguari, G.M.Araújo 382 (NY). Paraíba, São João do Cariri, A.V.Lacerda 258, 536 (HUEFS). Pernambuco, Chapada do Araripe, G.Fotius 3737 (HUEFS). Piauí, Caracol, R.M.Santos 1400 (HUEFS). São Paulo, São Paulo, M.Koscinski s/n (IPA). Tocantins, Colméia, G.Pedralli 3315 (HUEFS); Santa Rosa do Tocantins, G.Pedralli 3354 (HUEFS).
Hymenaea courbaril var. subsessilis–BRAZIL, Amazonas, Manaus, W.A.Rodrigues 7906 (INPA). Mato Grosso, Aripuanã, M.Gomes 576 (INPA). Pará, Porto Trombetas, H.C. de Lima 6808 (RB); S.M. de Faria 1241 (RB).
Hymenaea courvaril var. villosa–BRAZIL, Paraíba, Near Paquivera de Netuba, Y-T.Lee 82 (NY).