





Citation: Zuntini AR, Lohmann LG (2014) Synopsis of Martinella Baill. (Bignonieae, Bignoniaceae), with the description of a new species from the Atlantic Forest of Brazil. PhytoKeys 37: 15–24. doi: 10.3897/phytokeys.37.6940
Martinella has traditionally included two species, Martinella iquitoensis and Martinella obovata, that are characterized by the presence of interpetiolar ridges surrounding the stems and minute prophylls of the axillary buds. A third species, Martinella insignis, is here described as new, illustrated and compared to other species in the genus. Martinella insignis is the first record of the genus in the Atlantic Forest of Brazil, and differs from other species of Martinella by the yellow corolla (vs. red to dark purple) and 5-lobed calices (vs. 2–4-lobed).
Martinella, Bignonieae, Bignoniaceae, Neotropics, Brazilian Atlantic Forest
Martinella Baill. (1888) is strongly supported as monophyletic by molecular data (

Morphology of Martinella insignis: A Habit B Node with interpetiolar ridge C Glandular stipitate trichomes D Flower (lateral view) E Calyx (opened) and gynoecium F Fruit G Root system with tuberous portions. From Zuntini 151 (A–E), Sucre 5519 (F) and Zuntini 321 (G).
The genus as currently circumscribed includes two species of neotropical lianas (
This work is based on the study of herbarium collections of Martinella deposited in CEPEC, MBM, MBML, MO, NY, RB, SPF and VIES (herbarium acronyms follow
Martinella martinii (DC.) Baill. ex K. Schum. (= Martinella obovata (Kunth) Bureau & K. Schum)
Roots with tuberous portions. Branches terete, glabrous or puberulous, with trichomes simple or stipitate-glandular, with continuous interpetiolar ridges, without interpetiolar glands; prophylls minute, triangular, glabrous or puberulous. Leaves 3-foliolate or 2-with the terminal leaflet modified into a simple or trifid tendril; leafets membranous to coriaceous, margins entire (sinuate), with or without mite-domatia, glabrous to puberulous, with glands on adaxial surface. Inflorescences axillary, arranged in racemes, panicles, thyrses or compound dichasia. Flowers with calyx tubular (campanulate), bilobed, irregularly 2–4-lobed, or 5-lobed, with lobes rounded or aristate, membranous, with few scattered glands; corolla deep purple, red or yellow, tubular in the basal portion and campanulate in the upper part, straight to weakly curved, membranous, glabrous outside, glabrous inside except with few glandular trichomes at stamen insertion; stamens included, glabrous, pollen in monads; ovary terete, smooth, glabrous or lepidote, with a single series of ovules on each placenta, style glabrous, stigma rhombic, glabrous. Capsules drying dark brown, linear, flattened, smooth, glabrous or puberulous, with calyx caducous; seeds oblong, winged, with wings opaque.
Martinella comprises three species, distributed from Mexico to eastern Brazil. The main features that distinguish the species are summarized in Table 1 and outlined in the key below.
Morphological and geographical summary of Martinella species, based on
| Character | Martinella insignis | Martinella iquitoensis | Martinella obovata |
|---|---|---|---|
| Leaflet texture | membranous | coriaceous | membranous to coriaceous |
| Leaflet shape | ovate | elliptic | ovate (elliptic) |
| Leaflet base | cordate | cuneate | cordate to truncate (cuneate) |
| Leaf domatia | absent | absent | |
| Tendril | trifid | simple (trifid) | trifid (simple) |
| Inflorescence | compound dichasium | thyrse or panicle | raceme |
| Calyx lobes | 5; aristate | 2–4; rounded | 2–4; rounded |
| Corolla color | yellow | dark purple | dark purple to red (lilac) |
| Distribution | Eastern coast of Brazil | Amazon basin | Antilles, Central America and Northern South America through Amazon basin |
| Soil preference | sandy | sandy | generalist |
| 1 | Calyx 5-lobed; corolla yellow; eastern Brazil | 1. Martinella insignis |
| 1’ | Calyx 2–4-lobed; corolla deep wine to red; Antilles, Central America and Northern South America through Amazon basin | 2 |
| 2 | Inflorescence arranged in thyrse or panicle; leaflet with cuneate base | 2. Martinella iquitoensis |
| 2’ | Inflorescence arranged in raceme; leaflet with cordate to truncate or rarely cuneate base | 3. Martinella obovata |
urn:lsid:ipni.org:names:77138471-1
http://species-id.net/wiki/Martinella_insignis
Figs 1–2BRAZIL. Bahia: Itamaraju, Rodovia Itamarajú-Teixeira de Freitas, 3km de Itamaraju (BR-101). Fazenda Chapadão, 3 November 1983, R. Callejas, A. M de Carvalho & L. M. Silva 1629 (holotype MBM-94960!; isotypes CEPEC not seen, MO- 3600686!, NY-00483568!, RB-232556!).
Martinella insignis differs from Martinella iquitoensis and Martinella obovata by its 5-lobed calyces and yellow corollas, in contrast to 2–4-lobed calyces and dark purple to red corollas in these other species (Table 1).
Lianas. Branches green, drying brownish, striated, densely covered with stipitate-glandular trichomes when young; prophylls 0.7–1.5 × 1.0 mm, densely covered with stipitate glandular trichomes, without patelliform glands (with few patelliform glands). Leaves 2-foliolate with the terminal leaflet generally modified into a trifid tendril; petioles terete, 34–64 mm long, covered with stipitate glandular trichomes; petiolules terete, 14–42 mm long, covered with stipitate glandular trichomes; leaflets weakly discolorous, membranous, ovate, with a long acuminate to caudate apex and a cordate base, margins entire (sinuate), 7.6–11.8 × 3.4–6.4 cm, glabrous except on margins and main veins of the abaxial surface where stipitate glandular trichomes are found, with pocket domatia on the axils of primary and secondary veins, with few glands concentrated near base and scattered along the mid-vein on the adaxial surface. Inflorescences compound dichasia, with up to 7 branching orders, 9.2–12.3 cm long, sparsely to densely covered with stipitate glandular trichomes; bracts linear to narrowly elliptic, 8.4–24.7 × 0.8–3.4 mm, densely covered with stipitate glandular trichomes; pedicels terete, 5.8–19.4 mm, sparsely to densely covered with stipitate glandular trichomes. Flowers with calyx pale green, tubular (campanulate), 8.8–15.4 × 6.5–12 mm, sparsely covered with stipitate glandular trichomes except densely covered at the base, with few glands near the apex; lobes 5, very shallowly triangular, aristate, aristae 2.9–8.6 mm long, densely covered with stipitate glandular trichomes; corolla yellow, weakly curved, 29.5–48.3 mm long, tubular basal portion 12.7–18.9 long × 2.4–4.5 mm wide, upper campanulate portion 15.0–23.4 long × 9.2–17.2 mm wide, lobes subcircular, 3.8–9.8 × 6.1–10.9 mm, with ciliate margins; stamens in two lengths, longer ones 11.4–17.3 mm, shorter ones 12.1–15.8 mm, thecae 2.4–3.0 mm, glabrous; staminode 1.1–3.6 mm, glabrous; gynoecium 29.2–39.6 mm long; ovary glabrous; style glabrous; stigma rhomboid, glabrous; nectariferous disk 2.3–3.0 × 1.0–1.8 mm. Capsules 33.6–73.0 × 1.1–1.2 cm, pubescent when immature, glabrous when developed. Seeds ca. 1.0 × 4.6 cm.

Details of Martinella insignis: A Flower B Leaflet base showing glands (abaxial face) C Mite-domatia between primary and secondary veins (abaxial side) D Root system with tuberous portions E Calyx detailing the aristae F Arista detail with glandular trichomes G Simple, tector trichomes on immature fruit. From Zuntini 151 (A–C, E, F), Zuntini 321 (D) and Demuner 4481 (G). Micrographs were obtained using focus stacking.
Martinella insignis is restricted to the northern portion of the Brazilian Atlantic Forest, occurring predominantly in areas with sandy soils (Fig. 3).
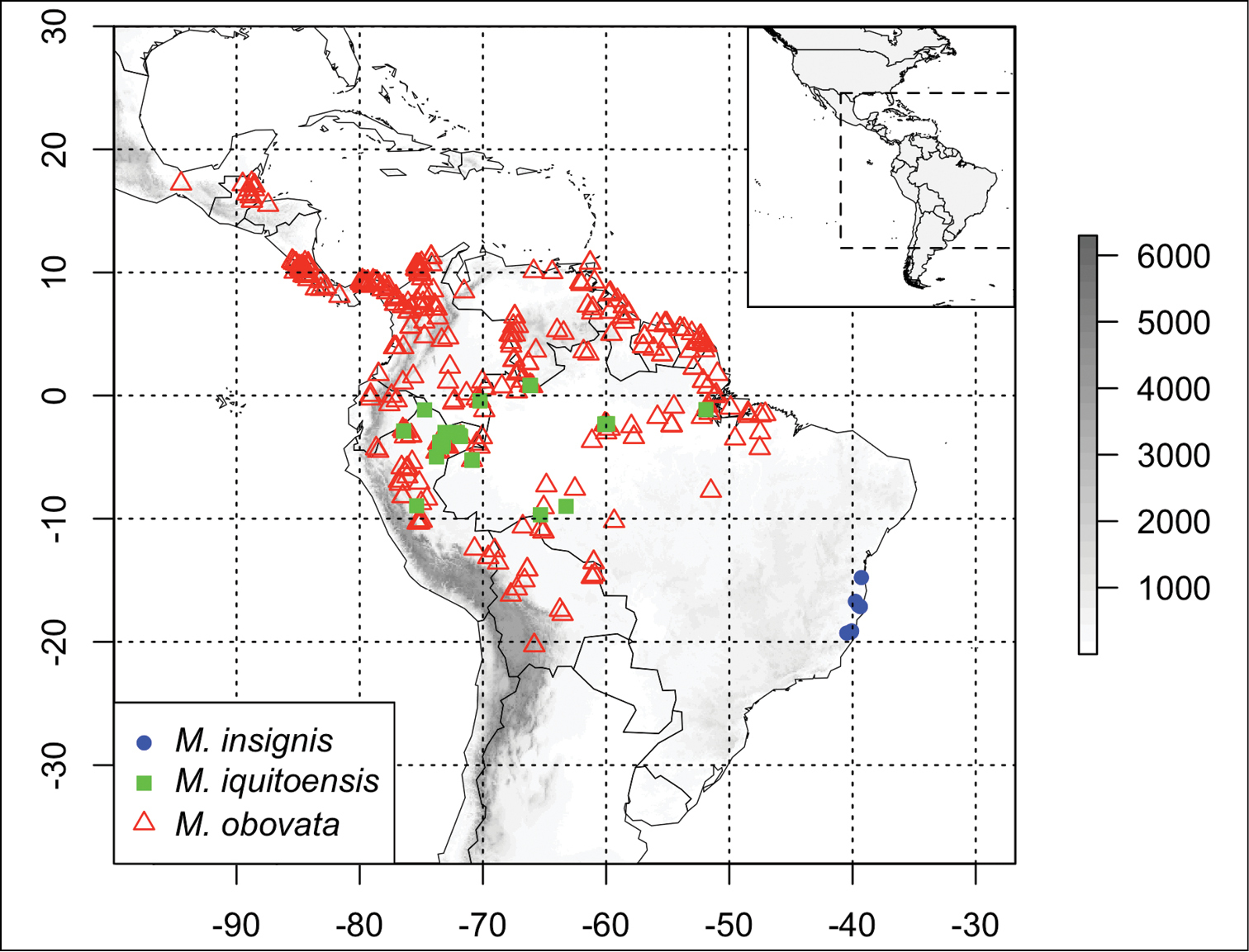
Distribution of Martinella species: Martinella insignis (solid blue circle), Martinella iquitoensis (solid green square) and Martinella obovata (open red triangles). Elevation in meters, according to the scale presented on the right.
The species epithet means remarkable or clearly distinguishable. This epithet was probably selected by Alwyn Gentry as reference to the contrasting floral color among species of Martinella.
Flowering specimens were collected between September and February and fruiting collections in January, September and November.
This species is considered Data Deficient [DD] according to
Martinella insignis is the first species of Martinella found in the Atlantic Forest of Brazil. This new species clearly belongs to Martinella based on its prophylls, the continuous interpetiolar ridges and the corolla shape. However, Martinella insignis can be distinguished by the membranous leaflets, 5-lobed aristate calyces, and yellow corollas. In addition, Martinella insignis also has pocket-shaped leaf domatia (Fig. 2C) and a puberulous indument of glandular stipitate trichomes that covers almost all organs, with variable density (Fig. 2E–F). These trichomes may also be found in Martinella obovata and a few other species in Bignonieae, and are typically formed by a multicellular secretory head, supported by a uniseriate stalk (
Brazil. Bahia: Guaratinga, Fazenda Vitória, 16°43’S, 39°46’W, 29 October 1979, L.A. Mattos Silva & H.S. Brito 634 (CEPEC, MO). Itabuna, Alcobaça para(ramal) S. Antonio, 24 January 1972, R.S. Pinheiro 1759 (CEPEC, MO). Itamaraju, S. Mori, L.A. Mattos Silva & T.S. Santos 10743 (CEPEC, MO), Itamaraju, Fazenda Riacho das Pedras, prop. Sr. Gersino Antônio Bronzon, 17°08'48"S, 39°21'53"W, 12 February 2007, R.A.X. Borges, A. Amorim, W.W. Thomas, L.C. Gomes, S. Sant’Ana & O. Cruz 825 (CEPEC, SPF). Espírito Santo: Linhares, Reserva Natural da Companhia Vale do Rio Doce (“Reserva de Linhares”), MME, 19°07'57.5"S, 40°04'06.3"W, 65m, 14 December 2007, A.R. Zuntini, W.A.A. Pires & G.S. Siqueira 151 (CVRD, RB, SPF), A.R. Zuntini, E. Françoso, J. Lopes & V. Augusto 321 (SPF). Governador Lindenberg, Pedra de Santa Luzia, 420–590 m, 7 November 2007, V. Demuner, T.A. Cruz & R.R. Vervloet 4481 (MBML, SPF). Sooretama, Mata de tabuleiro situada ao Noroeste da sede da Reserva da Sooretama, 14 July 1969, D. Sucre 5519 (RB–photo).
http://species-id.net/wiki/Martinella_iquitoensis
PERU. Loreto: Iquitos, 23 February 1924, J.G. Kuhlmann 1492 (holotype RB-22027!; isotypes, MO-2192060!, RB-22027 [second sheet]!).
This species is distributed widely in the Amazon basin (Brazil, Colombia, Ecuador, Peru and Venezuela), typically in sandy soils (
Martinella iquitoensis is distributed geographically through an area that is < 2000 km2, with seven
This species was distinguished by Sampaio from Martinella obovata based on the corolla color and size, leaflet texture and size, tendril type, and calyx indument. However these characters have proven to be fairly variable, especially in Martinella obovata, leading to morphological overlap between those taxa. The lack of a clear morphological discontinuity combined with the sympatric distributions, make these species hard to separate. Moreover, the difficulty in delimitating these two species can be observed in the few treatments that dealt with those species, which is particularly evident in the contrasting species keys presented (
A character that might help telling these species apart is the inflorescence structure: a thyrse or panicle in Martinella iquitoensis versus a raceme in Martinella obovata. This character, combined with leaflet base, is here proposed as diagnostic for each species; however, the examination of additional material is necessary to validate its usefulness and consistency.
http://species-id.net/wiki/Martinella_obovata
COLOMBIA. Turbaco, s.d., Humboldt and Bonpland 1391 (holotype, P-Bonpl. [P00670823]!)
This species is found from Central America and the Antilles through northern South America to the southern Amazon basin, in the Antilles, Belize, Bolivia, Brazil, Colombia, Costa Rica, Ecuador, French Guiana, Guatemala, Guyana, Honduras, Mexico, Panama, Peru, Surinam, and Venezuela, in different soil types and habitats (
Martinella obovata is distributed geographically through an area that is ≥ 2000 km2, with < 20% of its known individuals occurring outside Protected Areas, and seventy-two
This species is the most variable and has the largest distribution in the genus (Central America and Caribbean throughout Amazon basin). Such variation is product of its phenotypic plasticity and wide ecological range, and is responsible for making this species hard to be distinguished from Martinella iquitoensis (as discussed above). Nonetheless, this high variation may also be seen as an evidence of a species complex, in which case, additional studies would be needed. For a complete list of synonyms see
We thank CNPq for an undergraduate fellowship to ARZ (113717/2006-7) and a Pq-1 grant to LGL; FAPESP for a research grant to LGL (2011/50859-2), two FAPESP/BIOTA-NSF/Dimensions of Biodiversity Grants (2012/50260-6 and 2013/50297-0), and Vale S.A. for financial and logistic support. We also thank Dr. Charlotte Taylor for her review and significant comments on this manuscript; Iván Jiménez, Seema Sheth and Trisha Consiglio for assistance with the establishment of the conservation status of taxa; the editor and anonymous reviewers for their constructive remarks; Dr. Rafaela Forzza (RB) and Dr. Adriana Lobão (UFF) for providing images of specimens; Klei Sousa for the illustration and Geovane Siqueira for assistance during field work.