





Citation: Bellot S, Renner SS (2014) The systematics of the worldwide endoparasite family Apodanthaceae (Cucurbitales), with a key, a map, and color photos of most species. PhytoKeys 36: 41–57. doi: 10.3897/phytokeys.36.7385
Using morphological, nuclear, and mitochondrial data, we here revise the taxonomy of Apodanthaceae and allocate the 36 names published in the family to ten biological species in two genera, Apodanthes and Pilostyles. All species are endo-parasites that live permanently inside trees or shrubs of the families Salicaceae or Fabaceae and that only emerge to flower. Because of this life history, Apodanthaceae are among the least known families of flowering plants. Nevertheless, the World’s herbaria as of 2013 hold at least 785 collections that, in combination with DNA phylogenies, permit well-founded species circumscriptions and geographic range maps. We also provide a key to all species, discuss the newly accepted or synonymized names, and make available color photos of six of the ten species.
Apodanthaceae, genus circumscriptions, mitochondrial DNA sequences, nuclear DNA sequences, parasitic plants, species circumscriptions
Apodanthaceae Tiegh. ex Takht. (Cucurbitales) is a family of endoparasites that live entirely in their host’s stems and only become visible once the strictly unisexual flowers have burst through the bark. This life style, added to the small size of the flowers and patchy occurrence of the apparently mostly dioecious populations, has made it difficult to collect good and complete herbarium material (including both sexes and flowering and fruiting specimens). While populations once identified may be recollected at the same time year after year, usually only local botanists will have the opportunity to carry out such recollections. Apodanthaceae are disjunctly distributed in North and South America, mainland Africa, Iran, and Australia. They occur in arid as well as humid tropical environments. Two genera have been validly described, the worldwide Pilostyles, and Apodanthes from Central and South America.
The taxonomy of the genera and species of Apodanthaceae
has not been studied since the work of Ida de Vattimo-Gil
(
To achieve a better understanding of species boundaries
and relationships, and to clarify the species’ geographic
and host ranges, we compared loans from numerous herbaria,
dissected flowers, and isolated DNA from multiple
collections. Molecular markers useful for these obligatory
holoparasites are the nuclear 18S ribosomal RNA region and
mitochondrial matR (
We enlarged the DNA data matrix of
Total genomic DNA was extracted from herbarium specimens
using the commercial plant DNA extraction Invisorb® Spin
Plant Mini Kit (Stratec molecular, Berlin, Germany). The
mitochondrial matR and the nuclear 18S genes were
amplified using the primers listed in
We geo-referenced locality data from 785 herbarium collections on loan from the herbaria B, G, C, GH, K, M, MO, MSB, W, NA, PMA, and SI and added data from the Global Biodiversity Information Facility (GBIF Backbone Taxonomy, 2013-07-01, http://www.gbif.org/species/7279680). We also recorded host names, up-dating their taxonomy as relevant. All label information was compiled in a database using the Botanical Research and Herbarium Management System (BRAHMS, http://herbaria.plants.ox.ac.uk/), and maps were produced using DIVA-GIS 7.5 (http://www.diva-gis.org). Collections were sorted by geography, flowering specimens were sexed to evaluate sexual dimorphism, and a representative number of flowers were then dissected under a stereoscope. For each dissected flower, the first author recorded the number, arrangement and size of the tepals, shape and ornament of the pistil/central column, number of pollen sacs, presence of hairs and presence of a nectary at the base of the flower. Pictures of representative organs were taken using a Dino-Lite USB microscope model AM413ZT (Dino-Lite Europe) and the DinoCapture Imaging software version 2.0 of the same company.
The dissections showed that species have characteristic flower sizes, number of tepals, tepal cilia, and number of anthers rings. For the American species, we use these differences in the key (below). Suppl. material 2 shows measurements and counts from the 123 dissected flowers. Six collections could not reliably be assigned to these groups because their flowers were slightly unusual: R. Callejas et al. 8062, a male plant from Colombia identified as Apodanthes caseariae by A. Idarraga in 2002; Y. Mexia 4540, a female plant from Brazil that is the type of the name Apodanthes minarum; H. S. Irwin et al. 20350, a female plant from Brazil identified as Pilostyles ulei by Ida de Vattimo in 1975; H.S. Irwin 31560, a male plant identified as Pilostyles blanchetii by the first author but parasitizing an uncommon host (Dioclea, Fabaceae); J. Rzedowski 11303, a female plant from Mexico identified by the collector as Pilostyles thurberi; and F. Chiang 9034, a female plant from Mexico identified as Pilostyles thurberi by J. Henrickson in 1972.
The 18S and matR molecular trees show the Pilostyles collections that we wanted to identify (in red on Fig. 1) grouped with Pilostyles thurberi or Pilostyles blanchetii. The collections R. Callejas et al. 8062 and Y. Mexia 4540 grouped with two undoubted representatives of Apodanthes caseariae. R. Callejas et al. 8062 is a male plant and comes from the border with Panama, a country where Apodanthes caseariae has been repeatedly collected. The host of R. Callejas et al. 8062 was originally identified as Trema (Cannabaceae), but a partial matR sequence of this host BLASTed to Casearia nitida, making it likely that the host was in fact a Casearia. If that is the case, this would suggest that the collection represents an Apodanthes. The few male flowers of Apodanthes caseariae that have so far been dissected (Suppl. material 2) do not allow assessing the full morphological variability of the male flowers of this species. Therefore we had to rely on DNA for identification. In terms of its matR (Fig. 1A) R. Callejas et al. 8062 was embedded among other sequences of Apodanthes caseariae, while in terms of its 18S (Fig. 1B), it was sister to them. We identified the specimen as Apodanthes caseariae. Other matR and 18S sequences in the Apodanthes caseariae clade are from the type of the name Apodanthes minarum (Mexia 4540) from Brazil. Its host was a Casearia and its (female) flowers match those of Apodanthes caseariae (Suppl. material 2). We therefore synonymize Apodanthes minarum under Apodanthes caseariae (an action carried out below).
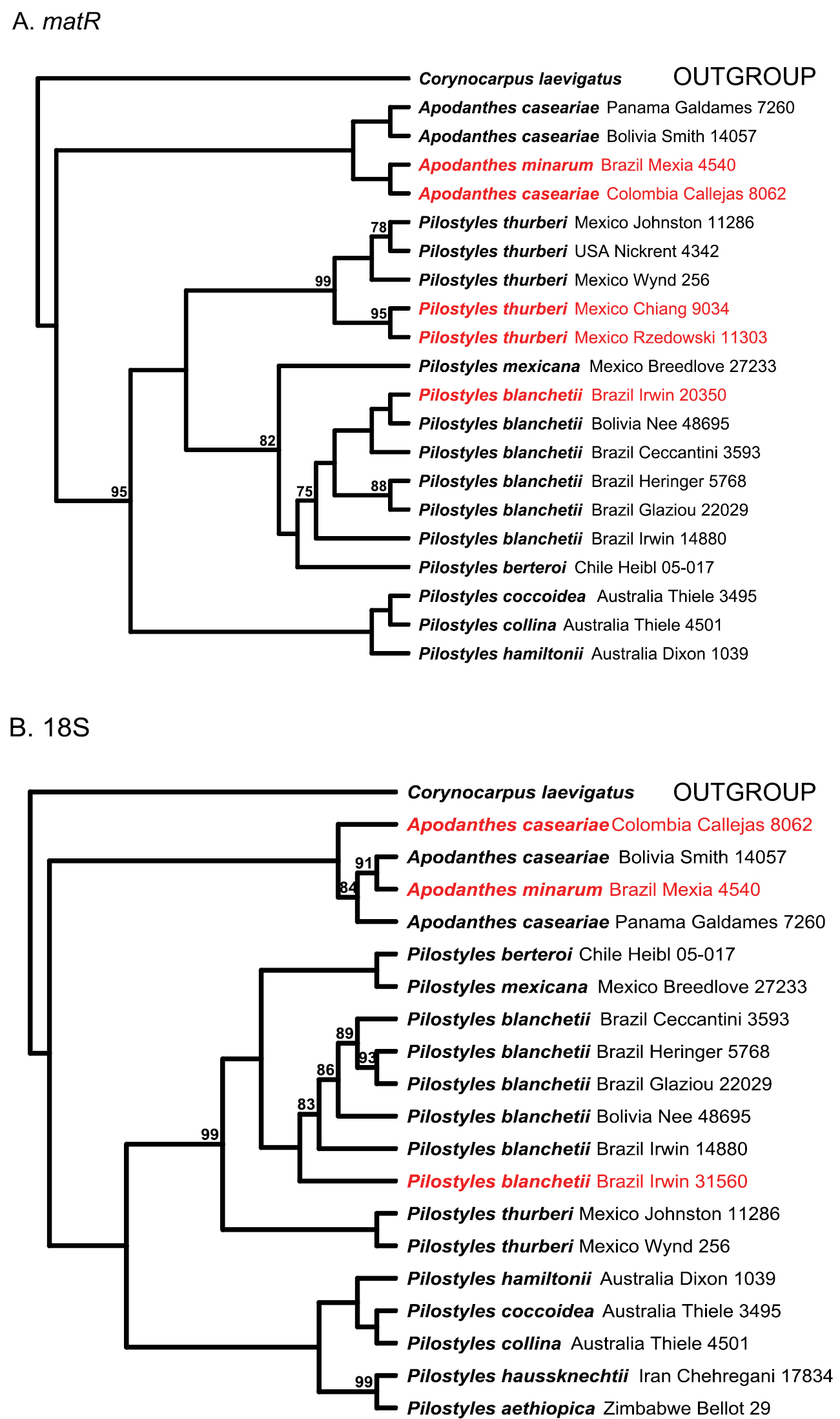
Phylogenetic relationships in Apodanthaceae obtained under maximum likelihood from the mitochondrial gene matR (A) and the nuclear ribosomal RNA gene 18S (B). Shown in red are the specimens we wanted to identify to species. Numbers indicate bootstrap support >70%.
In combination, the present morphological and molecular results show that Apodanthaceae comprise at least ten biological species that can be allocated to two mutually monophyletic genera. In the Americas, these are Pilostyles thurberi in the southern United States of America and Mexico, Pilostyles mexicana in Mexico, Guatemala and Honduras, the widely distributed Pilostyles blanchetii from Panama to Jamaica to Brazil and Uruguay, and Pilostyles berteroi in Chile and Argentina. The Americas also harbor Apodanthes caseariae from Guatemala to Brazil (Fig. 2). Australia has three species, Pilostyles coccoidea, Pilostyles collina, and Pilostyles hamiltonii; Iran has Pilostyles haussknechtii, and Africa has Pilostyles aethiopica. The second African species, Pilostyles holtzii has not been recollected since 1907, when its type collection was made. Another species, the southern Brazilian Pilostyles stawiarskii, is only known from two specimens (one of them the type) collected at the same locality in Jan./Feb. 1948 and Dec. 1949; morphologically it resembles Pilostyles blanchetii (Vattimo, 1950). The host ranges of our accepted genera and species do not overlap. Apodanthes parasitizes only Salicaceae, whereas Pilostyles parasitizes only Fabaceae. As seen on Figure 3, there is a correspondence, although not perfect, between the phylogenies of host genera and parasitic species, and host specialization may have played a role in speciation of Apodanthaceae. At the species level, Table 1 shows that species of Apodanthaceae can grow on one or up to thirteen host species. As seen in Figures 2 and 3, our species concepts are corroborated by geographic and host ranges, except in the case of Apodanthes caseariae and Pilostyles blanchetii, both widespread in Brazil. These two species have different sized flowers (see below), and parasitize phylogenetically distantly related hosts (Fig. 3).
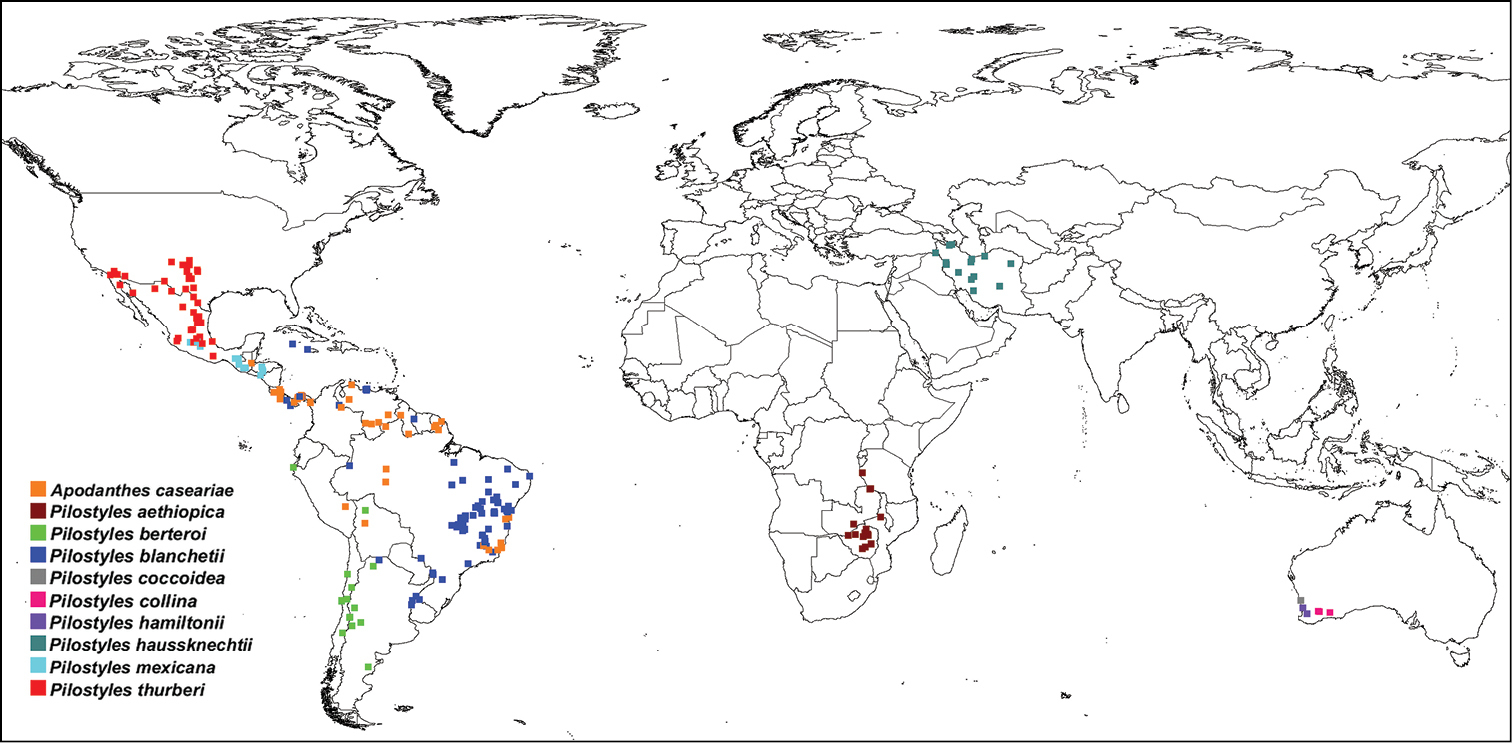
Geographic distribution of Apodanthaceae based on label information from 785 herbarium collections.
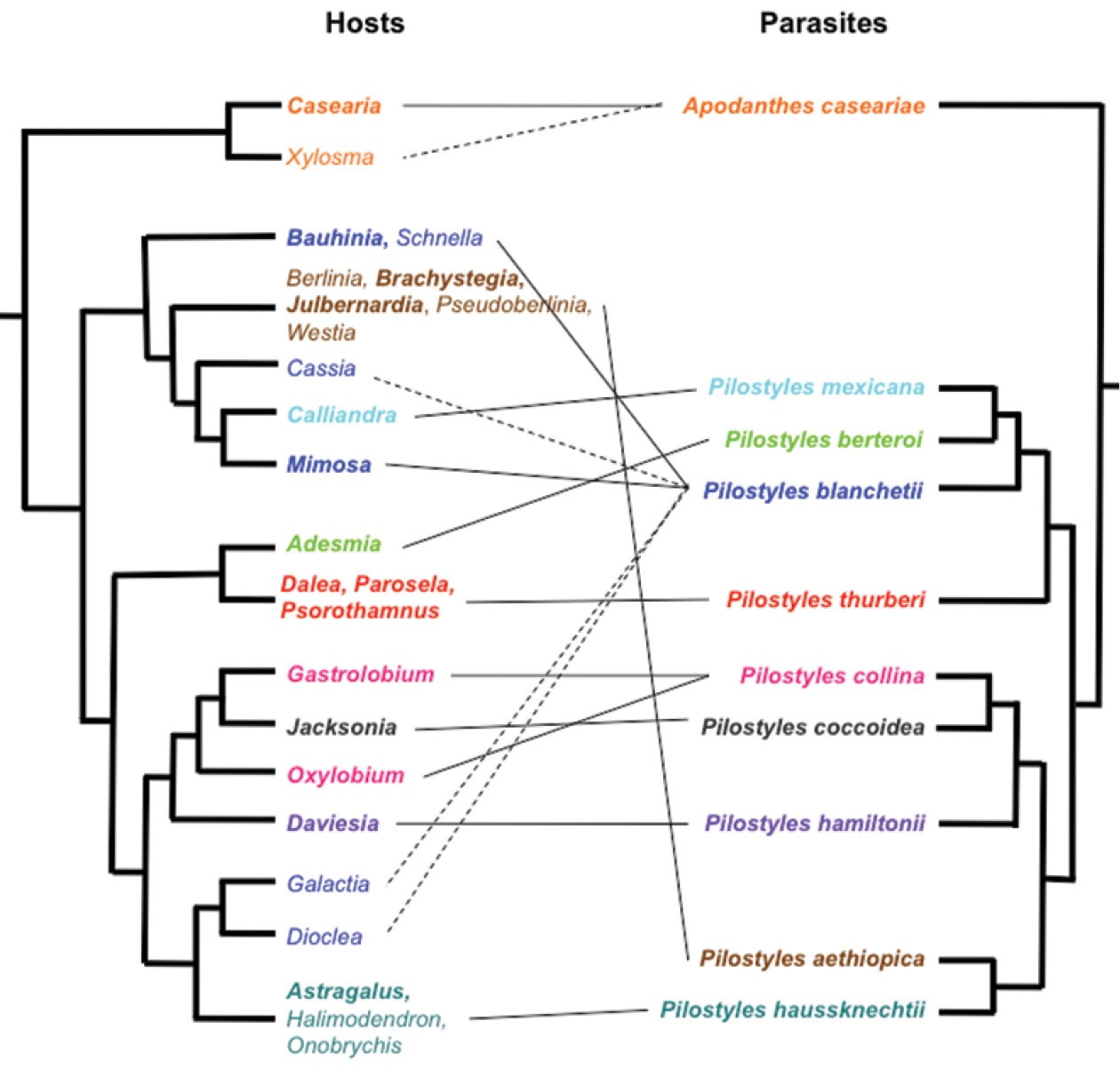
Phylogenetic relationships among the hosts of Apodanthaceae
(legume relationships from
Hosts of Apodanthaceae based on label information from 785 herbarium collections. Upper case numbers refer to the references below the table.
| Parasite | Host genera | Host species |
|---|---|---|
| Pilostyles aethiopica | Berlinia1, Brachystegia, Julbernardia, Pseudoberlinia1, Westia1 | Brachystegia boehmii Taub., Brachystegia glaucescens x spiciformis, Brachystegia spiciformis Benth., Brachystegia taxifolia Harms., Julbernardia globiflora (Benth.) Troupin |
| Pilostyles berteroi | Adesmia | Adesmia arborea Bert. ex Savi, Adesmia aff. spinosissima Meyen, Adesmia obovata Clos, Adesmia bedwellii Skottsb., Adesmia miraflorensis Remy, Adesmia uspallatensis Gill ex H. & A., Adesmia gracilis Meyen ex Vogel, Adesmia microphylla H. & A., Adesmia monosperma Clos, Adesmia pinifolia Gillies, Adesmia trijuga Gillies |
| Pilostyles blanchetii | Bauhinia, Cassia, Dioclea, Galactia2, Mimosa, Schnella | Bauhinia candicans Benth., Bauhinia divaricata L., Mimosa claussenii Benth., Mimosa cyclophylla Taub., Mimosa aff. setosa Benth., Mimosa maguirei Barneby, Mimosa scabrella Benth., Mimosa setosissima Taub., Mimosa uraguensis H. & A., Mimosa cf. xanthocentra Martius, Schnella cumanensis Britton & Rose |
| Apodanthes caseariae | Casearia, Xylosma | Casearia aculeate Jacq., Casearia arborea Urb., Casearia decandra Jacq., Casearia grandiflora Cambessèdes, Casearia guianensis Urb., Casearia hirsute Swartz, Casearia nitida Jacq. |
| Pilostyles coccoidea | Jacksonia | |
| Pilostyles collina | Gastrolobium, Oxylobium | Gastrolobium euryphyllum Chandler & Crisp |
| Pilostyles hamiltonii | Daviesia | Daviesia angulata Benth., Daviesia decurrens Meissner, Daviesia pectinata Meissner, Daviesia preissii Lindley |
| Pilostyles haussknechtii | Astragalus, Halimodendron, Onobrychis | Astragalus brachycalyx Fisch., Astragalus brachystachys DC., Astragalus cephalanthus DC., Astragalus chalaranthus Boiss. & Hausskn., Astragalus compactus Reiche, Astragalus floccosus Boiss., Astragalus gossypinus Fisch., Astragalus microcephalus Willd., Astragalus rhodosemius Boiss. & Hausskn., Astragalus spinosus Muschler, Astragalus susianus Boiss., Astragalus verus Olivier, Halimodendron halodendron (Pall.) Druce |
| Pilostyles mexicana | Calliandra | Calliandra houstoniana (Miller) Standley |
| Pilostyles thurberi | Dalea, Psorothamnus, Parosela3 | Dalea bicolor Humb. & Bompl. in Willd., Dalea formosa Torrey, Dalea frutescens Gray, Psorothamnus emoryi (Gray) Rydberg |
Stem-endoparasites, non-photosynthetic. No leaves, stem or
roots, instead an endophytic system of cells inside the
stem parenchyma of the host, flowers bursting through the
host bark. Flowers unisexual, plants dioecious or
monoecious, a point still insufficiently known; flowers of
both sexes on the same host or not. Pollination by flies
and bees, possibly also wasps (
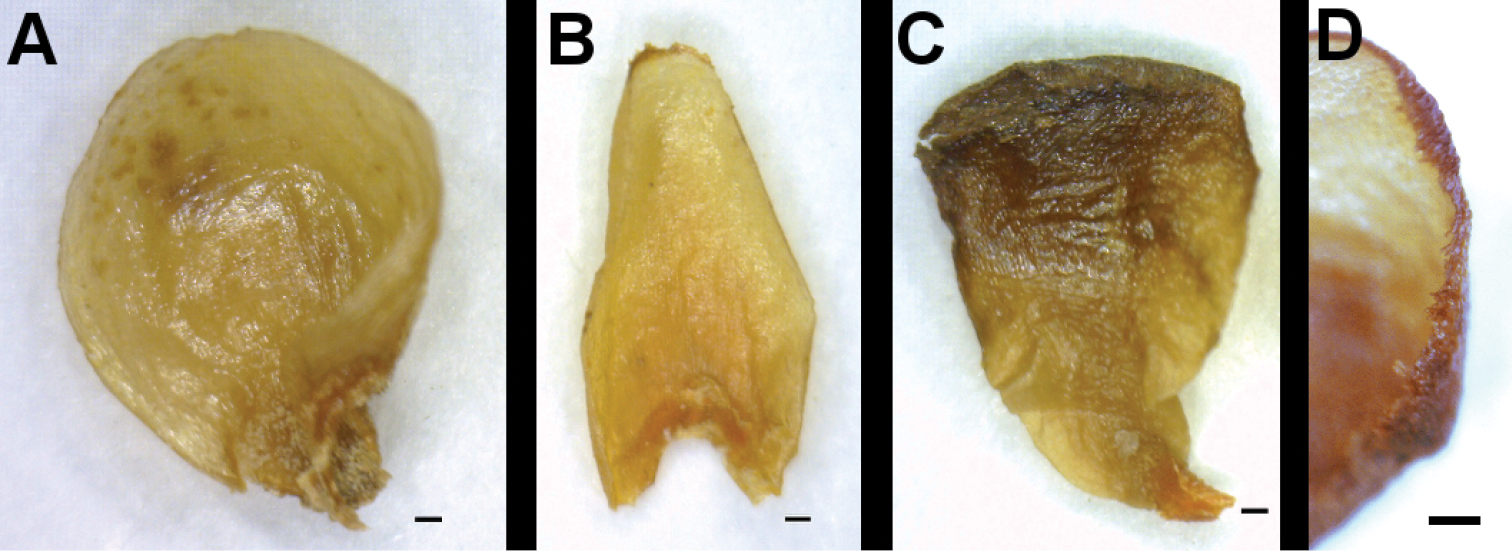
Tepals of Apodanthaceae. A Tepal of the outer whorl of Apodanthes caseariae B Tepal of the middle whorl of Apodanthes caseariae C Tepal of the inner whorl of Apodanthes caseariae D Tepal margin of Apodanthes caseariae. The scale bar corresponds to 0.2 mm.

Sexual organs of Apodanthaceae from rehydrated herbarium material. A Androecium of Pilostyles haussknechtii, note the two rings of pollen sacs topped by a ring of hairs B Style and stigma of Pilostyles haussknechtii C Ovary locule and ovules of Pilostyles haussknechtii D Androecium of Apodanthes caseariae after bursting of the pollen sacs, note the hairs covering the column apex E Gynoecium of Apodanthes caseariae. The scale bar corresponds to 0.2 mm.
| 1a | From the Neotropics, parasitizing Salicaceae, tepals always in 3 whorls with (from the outside) 2, 4, and 4 tepals, the inner whorl easily deciduous, female flowers >5 mm long | 1. Apodanthes caseariae |
| 1b | From the Neotropics, Africa, Iran, or Australia, parasitizing Fabaceae, number of outer tepals usually >2, female flowers <5 mm long | 2 |
| 2a | Occurring in Australia | 3 |
| 2b | Not in Australia | 5 |
| 3a | Tepals in 3 whorls | 2. Pilostyles collina |
| 3b | Tepals in 2 whorls | 4 |
| 4a | Flower diameter >3 mm | 3. Pilostyles hamiltonii |
| 4b | Flower diameter <3 mm | 4. Pilostyles coccoidea |
| 5a | Occurring in Africa | 5. Pilostyles aethiopica |
| 5b | Not in Africa | 6 |
| 6a | Occurring in Iran | 6. Pilostyles haussknechtii |
| 6b | Occurring in the America | 7 |
| 7a | Tepals in 3 whorls, each with 2 to 7 tepals, anthers in 4 whorls (spiral), number of anther lobes >70, on Adesmia | 7. Pilostyles berteroi |
| 7b | Tepals in 3 whorls, each with 3 or 4 (rarely more) tepals, anthers in 2 or 3 whorls, number of anther lobes <70, not on Adesmia | 8 |
| 8a | Middle tepals ovoid, on Calliandra, Dalea, Parosela or Psorothamnus, anthers in 3 whorls (at least if on Dalea, Parosela or Psorothamnus) | 9 |
| 8b | Middle tepals mostly diamond-shaped, apparently never on Calliandra, Dalea, Parosela or Psorothamnus, anthers in 2 whorls (females of the three species cannot be securely distinguished) | 8. Pilostyles blanchetii |
| 9a | On Calliandra, tepals in 3 whorls, each with 4 tepals | 9. Pilostyles mexicana |
| 9b | On Dalea, Parosela, or Psorothamnus, tepals in 3 whorls, each with 3 or 4 tepals. | 10. Pilostyles thurberi |
http://species-id.net/wiki/Apodanthes_caseariae
French Guiana, Karouany, c. 1802, parasitic on Casearia spec., P. A. Poiteau s.n. (P: P00686413).
Tepals white to yellow, female flowers >5 mm long, tepals in 3 whorls, the outer with 2 tepals, the inner one easily deciduous (Figs 4A–D; 6C, D). Growing in trunk and branches of Casearia and occasionally Xylosma (Salicaceae, Fig. 3) in Guatemala, Honduras, Costa-Rica, Panama, Colombia, Venezuela, Suriname, French Guiana, Brazil, Peru and Bolivia (Fig. 2).
Australia, Western Australia, Peak Charles, 10 Jan. 1982, parasitic on Oxylobium, B. D. Dell 8216 (G, MO).
Note: Tepals orange to red, in 3 whorls.
Growing in young stems of Gastrolobium
and Oxylobium
in Western Australia (Figs 2,
3,
see
Australia, Western Australia, Darling District, Helena Rover, Mundaring Weir, Mar. 1946, parasitic on Daviesia pectinata Lindl., C. D. Hamilton s.n. (PERTH, not seen).
Note: Tepals dark burgundy, in 2 whorls,
flowers >3 mm in diameter. Growing in young stems
of Daviesia
in Western Australia (Figs 2,
3,
see
Australia, Western Australia, Waddi Road, 30°33'26"S, 115°28'10"E, 7 Mar. 2008, parasitic on Jacksonia, K.R. Thiele 3495 (PERTH 07692447).
Tepals pale orange to brown, in 2 whorls, flowers
<3 mm in diameter. Growing in stems of Jacksonia
in Western Australia (Figs 2
and 3,
see
http://species-id.net/wiki/Pilostyles_aethiopica
Angola, Huila, 12 May 1860, parasitic on Berlinia paniculata Benth. = Pseudoberlinia paniculata (Benth.) P.A.Duvign., F. M. J. Welwitsch 529, 529b (C, G).
Tepals pink to brown, in 3 to 4 whorls each with 3-6
tepals. Male flowers with 1 or 2 ring(s) of ca. 15
pollen sacs, stamen filaments free from the central
column (Fig. 6F), hair cushion at the basis of the
inner tepals (

Flowers of Apodanthaceae species A Pilostyles berteroi on Adesmia in Chile (picture by C. Heibl) B Male flowers of P. thurberi on Psorothamnus emoryi in California (picture by L. Hendrickson) C Male and female flowers of Apodanthes caseariae on Casearia (?) in Panama (picture by G. Gerlach) D Close-up of a male flower of A. caseariae (picture by G. Gerlach) E Pilostyles blanchetii on Bauhinia in Panama; insert: close-up of a flower (pictures by C. Galdames) F Pilostyles aethiopica on Julbernardia globiflora in Zimbabwe (picture by S. Bellot); insert: close-up of male flowers (picture by D. Plowes) G Female and male flowers of P. haussknechtii on Astragalus in Iran (picture by S. Bellot). The white scale bars correspond to 1 mm and the black ones to 5 mm.
Middle East, parasitic on Astragalus, J. E. Haussknecht s.n (G-BOISS, not seen).
Note: Tepals pink to brown in 2 whorls, each with 6 to 10 tepals (Fig. 6G). Found at the basis of young branches of Astragalus and occasionally Onobrychis and Halimodendron in Iran (Figs 2, 3).
Chile, Quillota, parasitic on Adesmia, [in Chile 1828-1831] C. L. G. Bertero s.n. (P, not seen); Chile [from the collection number this was in “various localities in the Andes”, during the period from 27 Oct.-26 Nov. 1841], T. Bridges 1273 (BM, not seen, K, not seen).
Note: Tepals purple to brown with clearer margins (Fig. 6A), 9–18 in number, stamens in 4 whorls (spirals), with > 70 pollen sacs. Growing in older stems of Adesmia shrubs in Chile, Argentina, Peru, and Bolivia (Figs 2, 3). Our morphological (Suppl. material 2) and molecular data (Fig. 1) show that the species is nested among other species of Pilostyles, indicating that Gardner’s transfer was erroneous.
http://species-id.net/wiki/Pilostyles_blanchetii
Brazil: Bahia, 1839, J. S. Blanchet 2861 (NY).
Tepals purple to brown sometimes with clearer margins (Fig. 6E), in 3 whorls with usually 4 (rarely 3-6) tepals, the middle tepal diamond-shaped. Stamens in 2 whorls. Found in branches of Mimosa and Bauhinia, but also Cassia, Dioclea, Galactia and Schnella, in Jamaica, Cayman Islands, Costa-Rica, Panama, Colombia, Venezuela, Guyana, Brazil, Argentina and Uruguay (Figs 2, 3).
Mexico, near Zacuapan, Tenampa, parasite on Calliandra grandiflora Benth., Oct. 1906, C.A. Purpus 2207 (NY).
Tepals red to brown, in 3 whorls, each with 4 tepals. Growing in branches of Calliandra in Guatemala, Honduras and Mexico (possibly further south; Figs 2, 3).
http://species-id.net/wiki/Pilostyles_thurberi
USA, probably Arizona, near Gila river, June 1850, parasitic on Psorothamnus emoryi (A. Gray) Rydb., G. Thurber 682 (NY).
Tepals white, red to brown, in 3 whorls, each with 3 or 4 tepals, rarely more (Fig. 6B). Growing in branches of Dalea, Parosela and Psorothamnus in the southern United States of America and Mexico (Figs 2, 3). New York (NY) has a specimen from Mexico of this species annotated as “Pilostyles mortoni”, a nomen nudum, by Ida de Vattimo in 1952.
Flavio González and Natalia Pabón-Mora, at the university
of Antioquia in Colombia, are studying the ecology and
morphology of Apodanthaceae
in Colombia (
By combining morphological and molecular information, we show that Apodanthaceae comprise 10 species and that morphological distinctions fit well with geographical disjunctions and specializations on different hosts (Salicaceae vs. Fabaceae). DNA sequences of mitochondrial matR and nuclear 18S rDNA, along with morphology and geography permit identifying any collection of Apodanthaceae. A wider sampling of the morphological variation, especially of male Apodanthes caseariae and female Pilostyles blanchetii, Pilostyles mexicana and Pilostyles thurberi, however, is needed to determine whether some unusual specimens might deserve to be ranked as subspecies.
We thank C. Galdames, G. Gerlach, C. Heibl, L. Hendrickson, D. Plowes for photos of Apodanthaceae, K. R. Thiele for providing Australian silica-dried material, the reviewers N. Filipowicz and D. Nickrent for corrections, and the herbaria listed in Materials and Methods for providing loans. Funding came from the German Research Foundation (DFG RE 603/16-1).
List of accessions used in this study with author names for each species, herbarium vouchers, and GenBank accession numbers
Authors: Sidonie Bellot, Susanne S. Renner
Data type: species list
Copyright notice: This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.
Results of the dissections of 123 flowers from 82 specimens of Apodanthaceae
Authors: Sidonie Bellot, Susanne S. Renner
Data type: measurements
Copyright notice: This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.