





(C) 2014 Gilberto Morillo. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Morillo G, Krings A (2014) A new species and a new combination in Phaeostemma (Apocynaceae, Asclepiadoideae, Gonolobinae). PhytoKeys 33: 41–50. doi: 10.3897/phytokeys.33.6453
Phaeostemma surinamensis Morillo & Krings, sp. nov., anew species of Apocynaceae (Asclepiadoideae, Gonolobinae) is described and illustrated, and the new combination Phaeostemma fucata (Woodson) Morillo & Krings, comb. nov., is made. The new species, known only from a lowland wet forest of Suriname, seems to be closely related to Phaeostemma fucata, which is an endemic to Ptari-tepui, a sandstone mountain in the southeastern edge of the Venezuelan Guayana.
Climbing milkweeds, Matelea, Suriname, Venezuela
Phaeostemma E. Fourn. (Apocynaceae, Asclepiadoideae, Gonolobinae) is a South American genus of twining vines, distributed from southeastern Venezuela to Argentina. Heretofore, five names have been published in the genus — Phaeostemma brandonianum Silveira, Phaeostemma glaziovii E. Fourn., Phaeostemma grandifolia Rusby, Phaeostemma riedelii E. Fourn. and Phaeostemma tigrina Woodson—however, based on morphology, neither Phaeostemma grandifolia [syn. Matelea dasytricha (Schltr.) Woodson] nor Phaeostemma tigrina appear to belong to Phaeostemma. The latter species appear to belong to the Andean lineage Lachnostoma Kunth (

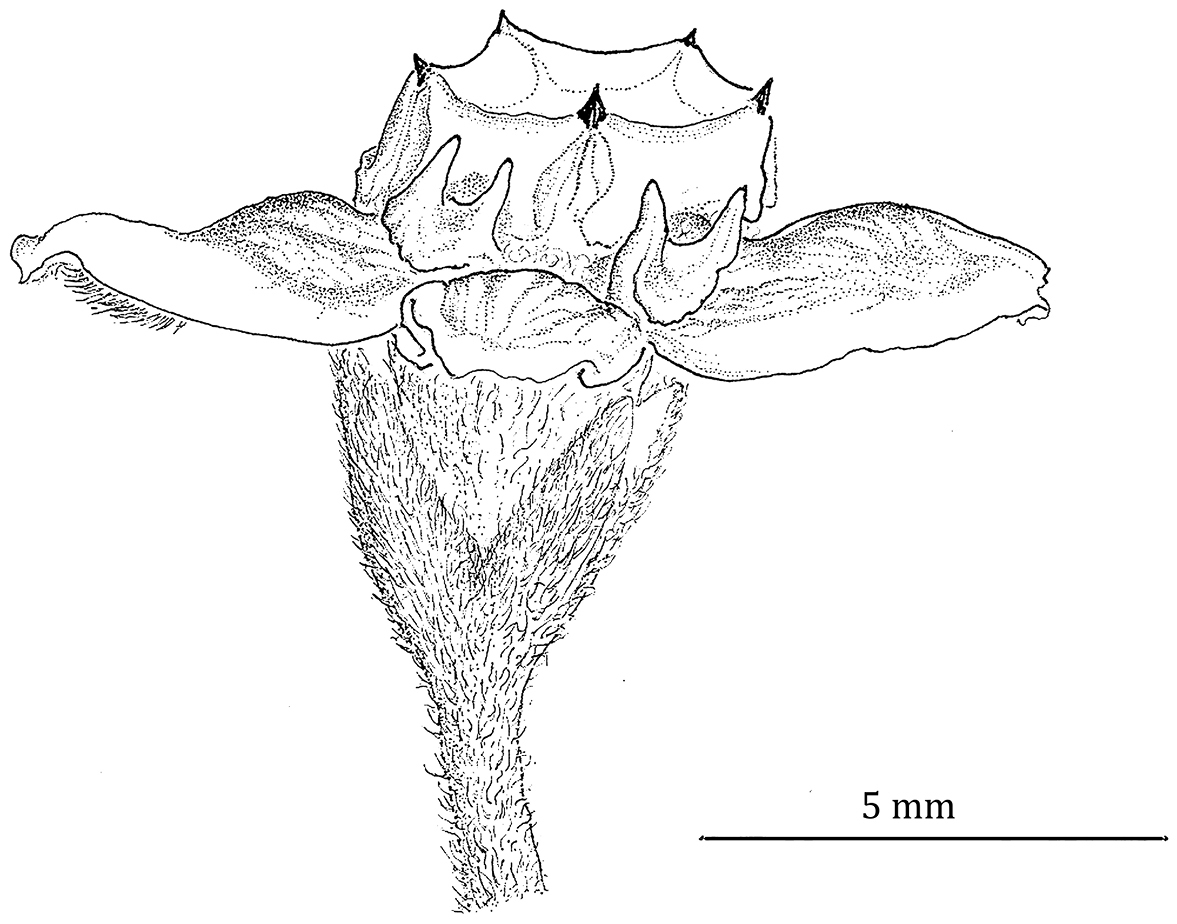
Flower of Phaeostemma fucata (from Steyermark 59963, MO).
Flower of Lachnostoma ecuadorensis Morillo (Homeier et al. 1174, QCNE).
Lachnostoma and Phaeostemma have been treated as nomenclatural synomyms of Matelea Aubl. sensu lato (
On-going work for various regional projects, including the Flora of the Guianas and the Biological Diversity of the Guiana Shield, has resulted in the discovery of a new species of Phaeostemma: Phaeostemma surinamensis Morillo & Krings [initially misidentified as Matelea glaziovii (E. Fourn.) Morillo = Phaeostemma glaziovii E. Fourn (
| 1a | Adaxial corolla lobe surface glabrous | 2 |
| 1b | Adaxial corolla lobe surface pubescent | 4 |
| 2a | Trichomes of stems, midveins, and inflorescences eglandular; corolla lobes longer than wide, adaxially obscurely reticulate when dry, leaf blades membranous; southeastern Brazil | Phaeostemma riedelii |
| 2b | Trichomes of stems, midveins, and inflorescences both eglandular and glandular; corolla lobes as long as wide or wider than long, adaxially conspicuously reticulate when dry, leaf blades membranous or coriaceous; Guayana and Suriname | 3 |
| 3a | Leaf blades coriaceous, broadly ovate to ovate-elliptic, marginally revolute, calyx lobes 8.5 mm long; Guayana (Venezuela) | Phaeostemma fucata |
| 3b | Leaf blades membranous, narrowly elliptic to oblanceolate-elliptic, marginally spreading; calyx lobes 5.5–6 mm long; Suriname | Phaeostemma surinamensis |
| 4a | Leaves subcoriaceous, veins on adaxial surface distinctly impressed, blades mostly broadly elliptic to ovate | Phaeostemma brandonianum |
| 4b | Leaves membranous, veins on adaxial surface not or only slightly impressed, blades mostly narrowly elliptic to slightly obovate | Phaeostemma glaziovii |
urn:lsid:ipni.org:names:77135536-1
http://species-id.net/wiki/Phaeostemma_fucata
Figure 1VENEZUELA. Bolivar, Ptari-tepui, densely forested slopes overlying sandstone, alt. 1800 m, 8 Nov 1944, J. Steyermark 59963 (Holotype: MO!; Isotypes: F!, NY!, VEN!).
urn:lsid:ipni.org:names:77135538-1
http://species-id.net/wiki/Phaeostemma_surinamensis
Figures 3–6A new species of Phaeostemma E. Fourn., morphologically similar to Phaeostemma fucata (Woodson) Morillo & Krings, but differing from the latter among other characters, by leaf blades thinner, membranous, narrowly elliptic to oblanceolate-elliptic, marginally spreading (vs. coriaceous, broadly ovate to ovate-elliptic, marginally revolute in Phaeostemma fucata), calyx lobes 5.5–6 mm long (vs. 8.5 mm long in Phaeostemma fucata), staminal corona lobes (Cs) 3–3.2 mm wide, narrower at lateral extremes (vs. 2.3 mm wide at apex, somewhat obtuse at lateral extremes [projections] in Phaeostemma fucata), and pollinia longer, 1–1.15 mm (vs. ca. 0.8 mm long in Phaeostemma fucata).
SURINAME. Lely Mts., SW plateaus covered by ferrobauxite, in secondary vegetation, at end of airstrip, alt. 550–710 m, 1 Oct 1975, J.C. Lindeman, A.L. Stoffers, A.R.A. Górts-van Rijn & M.J. Jansen-Jacobs 654 (Holotype: U!; Isotype: MO!).
Vine, woody, slender. Stems densely pubescent, pubescence mixed eglandular and glandular trichomes, eglandular trichomes ubiquitous reddish-brown, stiff, usually spreading, multiseptate, 1.2–2.3 mm long, glandular trichomes spreading, scarce in mature stems, 0.2–0.3 mm long. Leaf blades membranous, narrowly elliptic to oblanceolate-elliptic, 10.2–15.0 × 3.8–5.8 cm (1.6–1.8 cm wide at base), apex short acute, base narrowly and shortly cordate, trichomes of surfaces, veins, and margins mixed, yellowish-red, adaxial surface strigose, eglandular multicelled trichomes antrorse, curved to subappressed 0.7–1.3 mm long, glandular trichomes 3–4-celled, spreading, 0.15–0.2 mm long, present mainly on midvein, abaxial surface hispid or hispidulous, eglandular multicelled trichomes erect, 0.25–0.7 mm long, glandular trichomes spreading on the midvein, 0.1–0.2 mm long, midvein adaxially sulcate, abaxially prominent, lateral veins in 6–7 pairs, slightly to strongly prominent; colleters 2, digitate-cylindric, 2.8–3.3 mm long; petioles 1.1–1.7 cm long, densely pubescent, pubescence ubiquitous, eglandular multicelled trichomes spreading, 2.8–3.3 mm long, glandular trichomes 3–4-celled, spreading, 0.1–0.2 mm long. Inflorescence racemiform, 2–4-flowered, 1(–2) flowers open at a time; peduncles 3–5 mm long, sparsely to moderately pubescent, pubescence ubiquitous, eglandular trichomes spreading, 1.2–2 mm long, glandular trichomes 3–4-celled, scarce, spreading, 0.2–0.3 mm long, rachis with scars, 3–4 mm long, bracts oblong, 1.8–2 mm long, abaxially pubescent, eglandular trichomes multicelled, 0.3–0.4 mm long, pedicels 24–33 mm long, densely pubescent, pubescence ubiquitous, eglandular trichomes spreading, 0.9–2.5 mm long, glandular trichomes 3–4-celled, spreading, 0.15–0.3 mm long. Calyx 8.5 mm long, lobes green, dark red at apex, oblong-elliptic, 5.5–6 × 2.4–2.6 mm, apex obtuse, margins entire, adaxial surface mostly glabrous, except few trichomes at apex, abaxial surface densely pubescent, eglandular multicelled trichomes spreading or antrorse, 0.9–1.5 mm long, glandular trichomes 2–3-celled, spreading, 0.1–0.3 mm long, colleters 1 per sinus. Corolla pale green, with green vein network (fide collectoris), broadly subcampanulate, tube 5.5–5.8 mm long, up to 7 mm wide, sparsely short pubescent on the abaxial side, lobes imbricate in bud, deltoid, spreading, 8–9 × 9–10 mm, adaxial surface glabrous, abaxial surface densely pubescent, eglandular trichomes reddish-brown curved, antrorse, 0.5–1.5 mm long, glandular trichomes 3–4-celled, scarce, 0.15–0.25 mm long, apices obtuse-emarginate, margins entire. Gynostegium stipitate, style-head green yellow, somewhat concave, 4.3–4.5 mm in diameter, stipe 2.5 mm long, terminal style-head appendage absent. Corona gynostegial, fleshy, of 5 staminal (Cs) apically bifid segments fused to the corolla tube for most of its length, free in the upper 1.5–1.6 mm, apical area bifurcate, dorsally flattened, 3–3.2 mm wide at apex; anthers subtriangular, 1.9–2.1 mm wide between wings; nectar chambers ca. 2.5 × 2.6 mm. Pollinarium: corpuscula narrowly obovate-sagittate, ca. 0.5 × 0.33 mm long, caudicles ca. 0.22 mm long, pollinia triangular-pyriform, 1.0–1.15 × 0.6 mm. Follicles in very immature state, ca. 2 cm long, apparently 5-costate, densely glandular-pubescent, eglandular trichomes not seen, glandular trichomes 2–3-celled, 0.1–0.15 mm long. Seeds unknown.
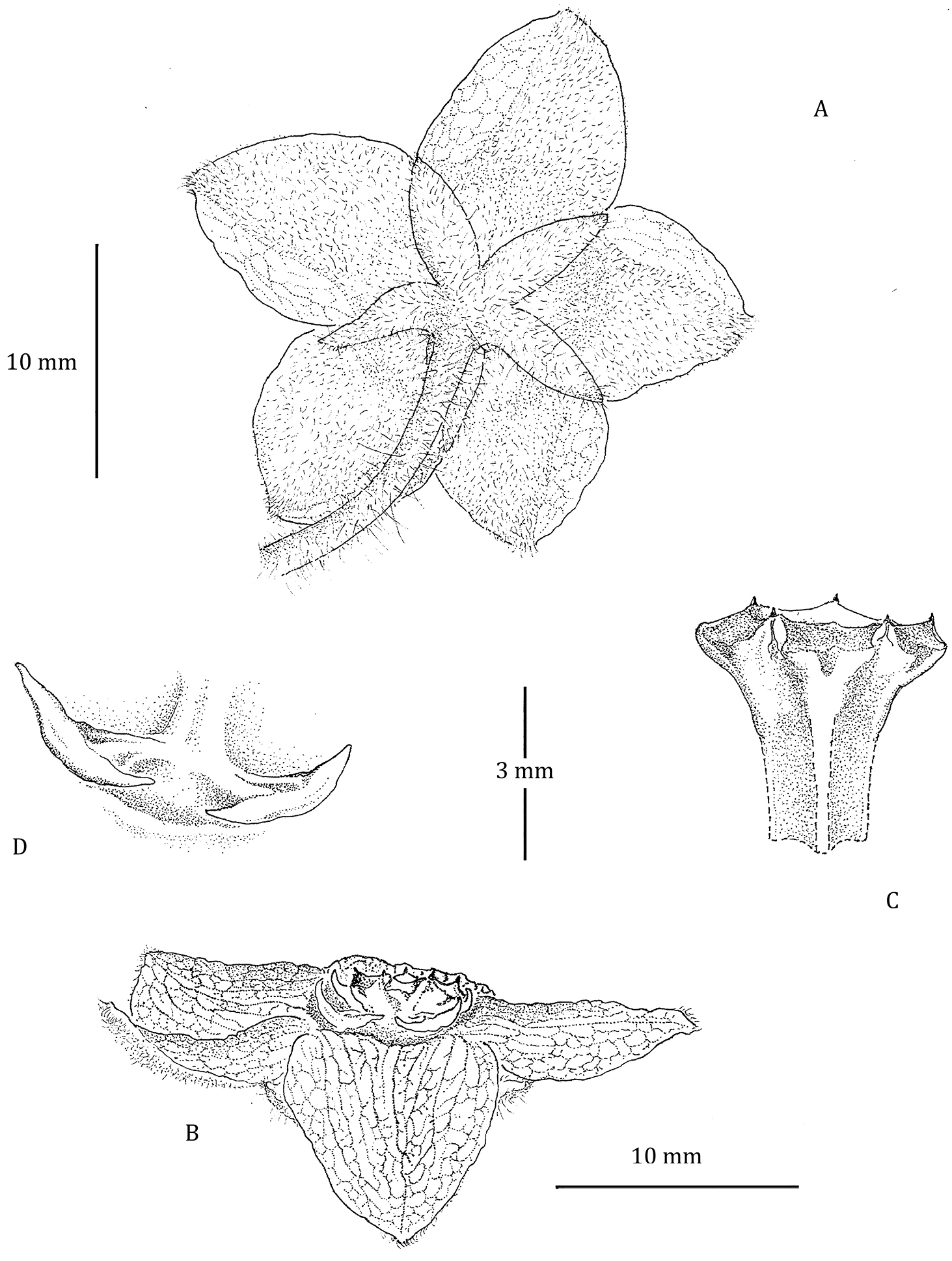
Flower of Phaeostemma surinamensis. A Abaxial view showing calyx and abaxial corolla lobes B Side view showing adaxial corolla surface and gynostegial corona C Style-head D Staminal corona (Cs). Based on Lindeman et al. 654 (U).

Holotype of Phaeostemma surinamensis (Lindeman et al. 654, U).

Detail of flower (side view) of Phaeostemma surinamensis from holotype (Lindeman et al. 654, U).

Detail of flower (adaxial view) of Phaeostemma surinamensis from holotype (Lindeman et al. 654, U).
Growing on a plateau covered with ferrobauxite rock. Endemic to Suriname, in tropical rain forests 500 to 710 m.
Collected in flower in October.
Currently, very little is known regarding the status of this species.
Phaeostemma grandifolia Rusby, Descr. S. Am. Pl. 101. 1920. = Matelea dasytricha (Schltr.) Fontella, Bradea 4 (9): 55. 1984 (syn. Gonolobus dasytrichus Schltr., Notizbl. Königl. Bot. Gart. Berlin 6(55): 177. 1914.)
Phaeostemma tigrina Woodson, Ann. Missouri Bot. Gard. 18: 560. 1931. = Lachnostoma tigrinum Kunth, Nov. Gen. Sp. (quarto ed.) 3: 199, t. 232. 1818[1819] (syn. Matelea humboldtiana Spellman & Morillo, Phytologia 34(2): 152. 1976.)
We thank the following herbaria for allowing the study of specimens pertinent to this study: MO, NY, U, US, VEN. We also thank the kind staff of MO and US for their hospitality during our recent visits.