





(C) 2013 Wendy A. Nelson. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Nelson WA, Dalen J, Neill KF (2013) Insights from natural history collections: analysing the New Zealand macroalgal flora using herbarium data. PhytoKeys 30: 1–21. doi: 10.3897/phytokeys.30.5889
Herbaria and natural history collections (NHC) are critical to the practice of taxonomy and have potential to serve as sources of data for biodiversity and conservation. They are the repositories of vital reference specimens, enabling species to be studied and their distribution in space and time to be documented and analysed, as well as enabling the development of hypotheses about species relationships. The herbarium of the Museum of New Zealand Te Papa Tongarewa (WELT) contains scientifically and historically significant marine macroalgal collections, including type specimens, primarily of New Zealand species, as well as valuable exsiccatae from New Zealand and Australia. The herbarium was initiated in 1865 with the establishment of the Colonial Museum and is the only herbarium in New Zealand where there has been consistent expert taxonomic attention to the macroalgae over the past 50 years. We examined 19, 422 records of marine macroalgae from around New Zealand collected over the past 164 years housed in WELT, assessing the records in terms of their spatial and temporal coverage as well as their uniqueness and abundance. The data provided an opportunity to review the state of knowledge of the New Zealand macroalgal flora reflected in the collections at WELT, to examine how knowledge of the macroalgal flora has been built over time in terms of the number of collections and the number of species recognised, and identify where there are gaps in the current collections as far as numbers of specimens per taxon, as well as with respect to geographical and seasonal coverage.
Herbarium, macroalgae, Museum of New Zealand Te Papa Tongarewa, regional floras
Herbaria and natural history collections (NHC) are critical to the practice of taxonomy. They are the repositories of vital reference specimens, enabling species to be studied and their distribution in space and time to be documented and analysed, as well as enabling the development of hypotheses about species relationships.
There are many challenges when using NHC for analyses of biota, particularly the potential sources of errors, accuracy and biases (
In terms of spatial and temporal data, older collections tend to be geo-referenced post-collection which may introduce location errors. The data associated with specimens are often highly variable as far as the level of detail provided, for example, the precision of the locality of the collection, habitat information, associated species, collection method, and whether multiple collections were made from within different habitats within a site. Some early collections (19th century, early 20th century) have only the year or month of collection provided with the specimen. Biases affect different aspects of the collections. Spatial biases can result from the position of access roads and settlements, particularly in the case of coastal collecting. Access to collection methods and equipment is also critical, for example, in the case of marine macroalgae in New Zealand there are large sections of coastline where access is only possible from the sea, and thus the use of boats is critical. Weather can have a significant impact on accessibility of sites and can lead to seasonal biases. For an important part of the flora sampling is only possible via SCUBA and via dredge equipment for deep-water samples. The location of active collectors has an impact on the number of collections obtained from particular regions, as does the perception of areas being of specific interest. Temporal biases can result from experts working actively on particular taxonomic groups, as well as from particular curatorial practices and personal interests (e.g. discarding damaged individuals, only accessioning a certain number of individuals) (
Collections result from targeted investigations as well as from opportunistic sampling. Sometimes remote areas are infrequently visited but have detailed and thorough collections because major effort is required to reach the area and very deliberate collections are undertaken. There are inevitably biases as far as which species are collected, with the potential for larger or more conspicuous species to be over-represented and with smaller or more difficult to collect species under-represented. In addition, sometimes common species are under-collected whereas rare or unusual species are collected more frequently.
In the mid to late 19th century there was a period of research on New Zealand algae by overseas research workers, particularly W.H. Harvey (Trinity College, Dublin), J.D. Hooker (Kew, London) and J.G. Agardh (Botanical Museum, Lund). In some cases material was deposited in New Zealand collections, e.g. collections made by William Colenso were sent to Hooker and Harvey and some material retained in New Zealand; collections made by Sven Berggren in 1874 were sent to Agardh with some duplicate material returned to New Zealand (
From the mid-1930s very significant collections were made by Victor Lindauer (
The major macroalgal collections are held in New Zealand in the herbaria of the Museum of New Zealand Te Papa Tongarewa (WELT), Landcare Research Manaaki Whenua (CHR) and at the Auckland Museum (AK) (
In the absence of a complete flora, there has been considerable recent effort directed to compiling and updating lists of currently accepted names and the taxonomic hierarchy, with published lists produced as part of the Species 2000 project documenting the New Zealand biota (
The WELT collections have been databased over a period of ca. 15 years. The recent focus within the herbarium has been on improving collection data and checking the dataset for errors, particularly grooming collection date data and mapping and verifying locality data. The collections at both AK and CHR have not been fully databased to date and have not received the level of scrutiny and identification that has been directed to the WELT collections. The AK and CHR collections currently do not have specialist marine phycologists associated with the collections. Because WELT collections have received expert identification and curation, they have been used as the primary source of data on the distributions of marine macroalgae for a number of research projects and government databases (e.g.
The grooming and updating of the database has provided an opportunity to review the state of knowledge of the New Zealand macroalgal flora reflected in the collections at WELT, and to ask a series of questions. Which regions of New Zealand are represented by the most comprehensive collections/least comprehensive collections? How has knowledge of the macroalgal flora built over time in terms of the number of collections and the number of species recognised? Are there patterns that can be discerned in the collection history and coverage? What proportion of the flora is represented by sufficient individuals for study and comparative investigation (number of specimens, geographic range, seasonal distribution)? Do these collections have the potential to enable other types of biodiversity analyses?
The data presented in this paper are drawn from the database of the Te Papa herbarium. Definitions for terms used in this paper are provided in Table 1. Data were assembled following several steps:
Definitions of terms used in this paper.
| Term | Definition |
|---|---|
| Collecting event | For each region collections were sorted by year, then collection date and precise location. Each unique combination of date and precise location was treated as a collection event. |
| Duplicate records | Specimen duplicates, i.e. same taxon with identical collection data. (Only 1 example of duplicate sheets (e.g. labelled a–c) was retained; duplicates with different registration numbers were removed.) |
| Season | Collection dates were grouped by month and allocated to seasons as follows: December-February = summer; March-May = autumn; June-August = winter; September-November = spring |
| Record | Single packet, box or specimen sheet |
| Taxon | Name used in database which includes identifications to family, genus and species level as well as tag names (informal names assigned usually in preliminary stages of investigations or for entities recognised in the field) |
Until the early-1990s, herbarium specimen data were available from the specimen labels and a hard copy register. The first electronic database system at Te Papa, Te Kahui, was custom-designed and implemented in 1993. Data were retrieved from the specimen labels and entered into the system by trained data-entry technicians. Where appropriate, extra information was sourced - the majority of this being latitude and longitude information derived from maps (NZMS 260 series). Most of the existing New Zealand algal specimens in the collection had an electronic record completed by ca. 2001. However, the record error rate was relatively high – in the order of 30-40%, with respect to coordinate data and locality information.
In 2005, all of the museum’s electronic records were migrated to an electronic collection information management system, KE EMu® (referred to as EMu), a relational database customised for museum collections. With the implementation of the new database, there was scope to improve the quality of information recorded. Features of EMu, such as sophisticated search functions and global updating options, have facilitated cross-checking for consistency in the locality records, use of place names and collector details. The verification and grooming of the algal data (e.g. cross checking of longitude and latitude, consistency of locality records, use of place names, collector details, identifying missing data fields and locating information where available) has become a core collection management activity since this time. However, much of this grooming effort has been somewhat opportunistic (for example, new acquisitions prompt a cross-checking of data for similar/nearby locations). Several special projects and requests for data have prompted more comprehensive data verification efforts. As part of this, several thousand backlog algal specimens were identified and databased; the database component of the work further prompted refinement to the consistency and accuracy of the locality data. Attention to the application of taxonomic concepts and names across the collection was also undertaken as part of this work.
In the absence of a published flora, a current species names list and taxonomic hierarchy is maintained on the Te Papa website (http://www.tepapa.govt.nz/ -
Figure 1 illustrates the regional boundaries applied in this exercise. The boundaries reflect a combination of biogeographic boundaries in previously published accounts of the marine biota (e.g.
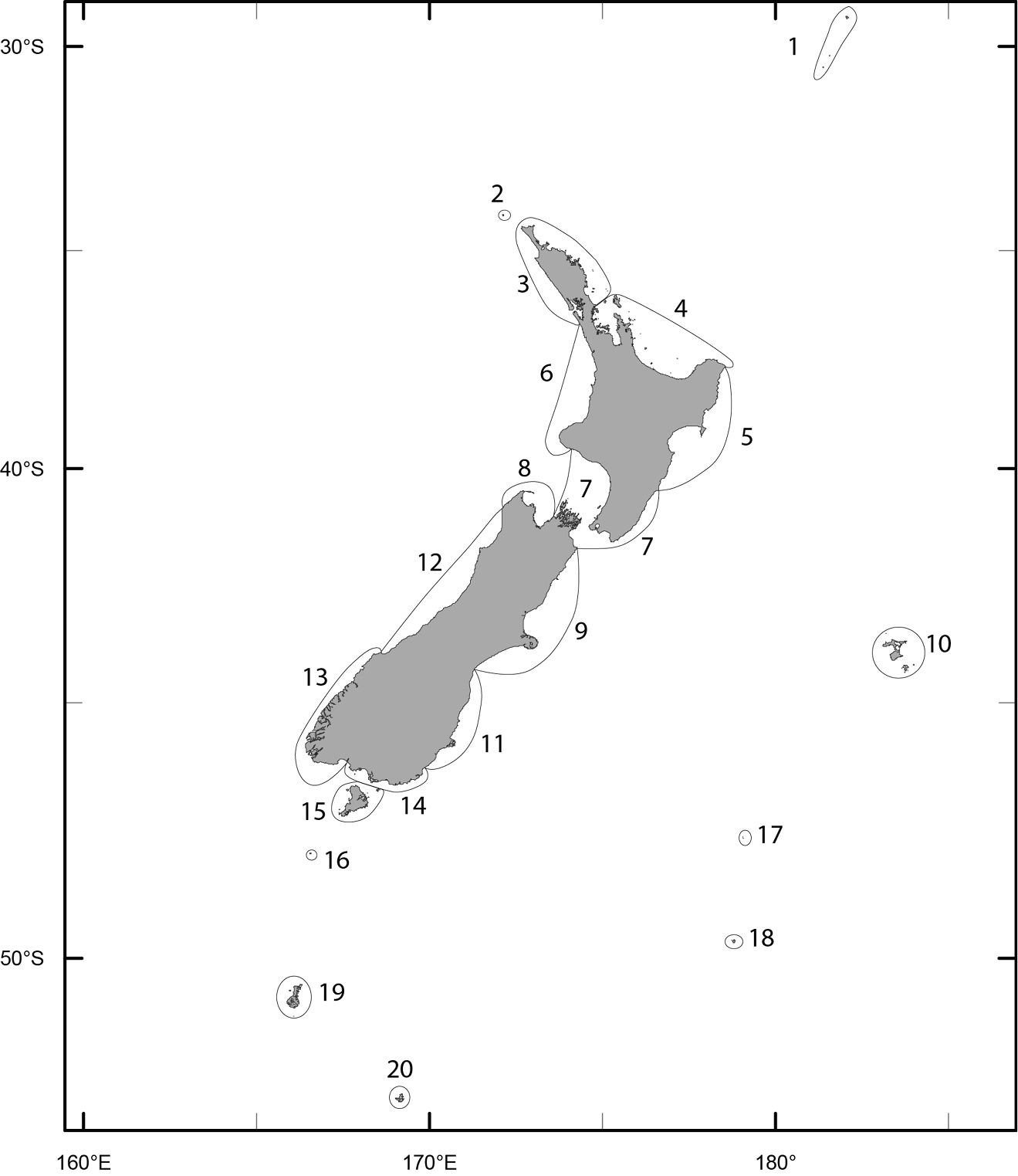
Map of New Zealand indicating the boundaries of the regions investigated in this study (1 Kermadec Is 2 Three Kings Is 3 North I. (NI) North 4 NI Bay of Plenty (BOP) 5 NI East 6 NI West 7 Wairarapa-Cook 8 South I. (SI) Northwest (NW) 9 SI Kaikoura 10 Chatham Is 11 SI Southeast (SE) 12 SI Westland 13 SI Fiordland 14 SI Southern 15 Stewart I. 16 Snares Is 17 Bounty Is 18 Antipodes Is 19 Auckland Is 20 Campbell I).
All New Zealand marine algal records were searched and grouped by latitude and longitude coordinates corresponding to the regions as defined above (20 categories) in Te Papa’s database. Records, current to December 2011, were exported by region into Excel spreadsheets. Table 2 summarises the data used in the analyses undertaken. The number of taxa includes all recognised entities present in the collections, including some that have been recognised as distinct at a family, genus or species level but are currently unnamed. It is important to note that the publicly accessible flora lists (
Summary of the specimen records and taxa analysed in dataset:
| Greens | Browns | Reds | Total | |
|---|---|---|---|---|
| Number of records | 2, 859 | 5, 495 | 17, 043 | 25, 397 |
| Number of unique records | 2, 213 | 4, 580 | 12, 629 | 19, 422 |
| Number of taxa | 118 | 199 | 679 | 996 |
| Number of classes | 3 | 3 | 4 | 10 |
| Number of orders | 7 | 13 | 20 | 40 |
| Number of families | 16 | 30 | 52 | 98 |
| Number of genera | 25 | 75 | 210 | 310 |
The number of new taxa represented in the collection by year of collection is presented in Figure 2, the cumulative total of taxa in the collections in Figure 3, and the number of taxa in the collections by decade of collection broken into divisions (red, brown, and green algae) in Figure 4. The surge in the number of collections around the 1870s reflects the material collected by both S. Berggren and H.H. Travers that was sent to Lund for examination by J.G. Agardh with duplicate material returned to New Zealand. Most specimens lodged prior to the 1930s were collected by W.A. Scarfe and R.M. Laing. In 1935 Josephine Tilden from the University of Minnesota, and a group of associates, visited New Zealand collecting in the Bay of Islands as well as on Stewart Island, and material was distributed as “South Pacific Plants”. V.W. Lindauer, the school teacher at Russell, Bay of Islands, was introduced to seaweeds by Tilden, resulting in his major contributions to New Zealand phycology (
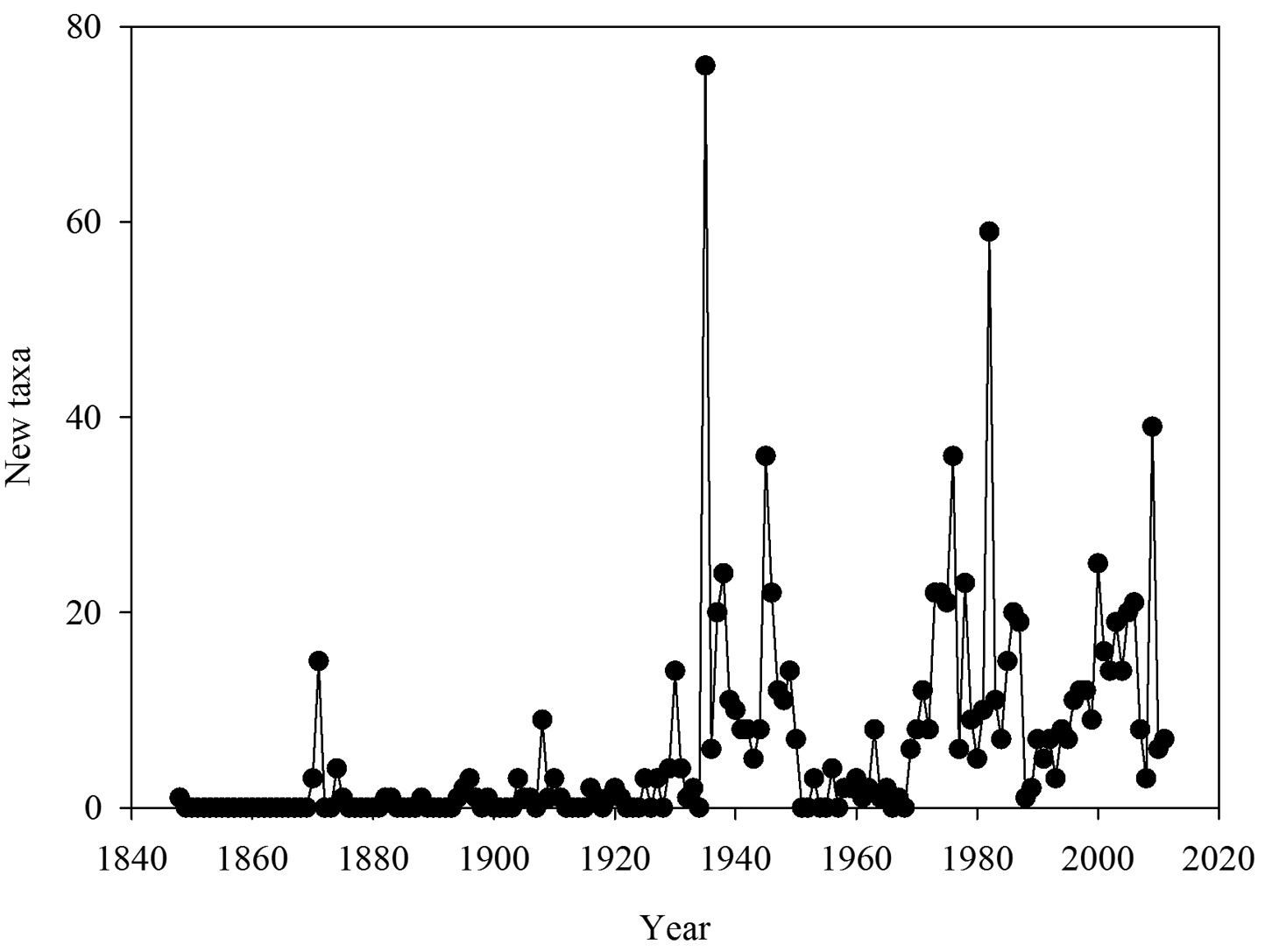
Number of new taxa represented in the collection by year of collection.
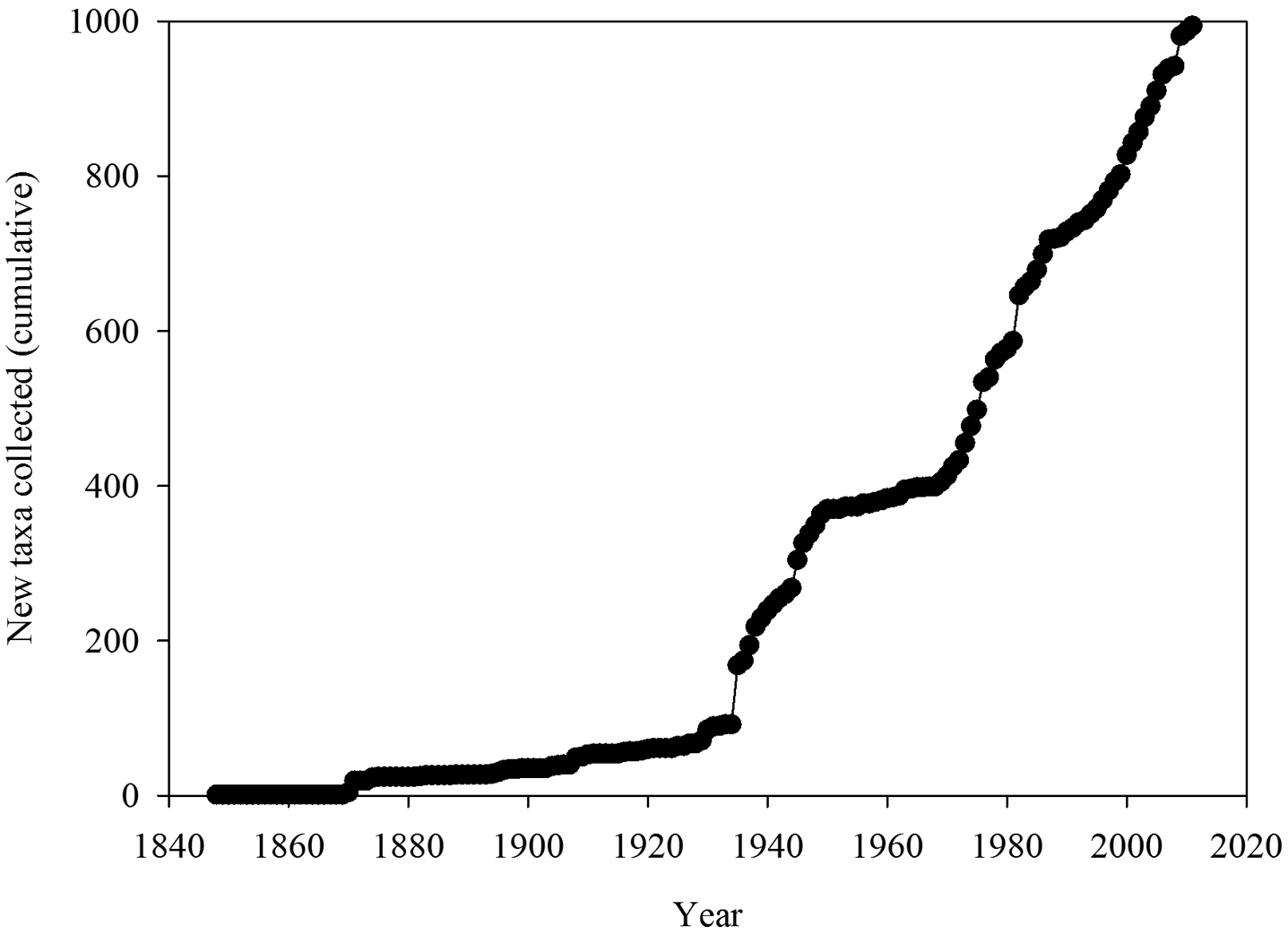
Cumulative total of taxa in the collections.
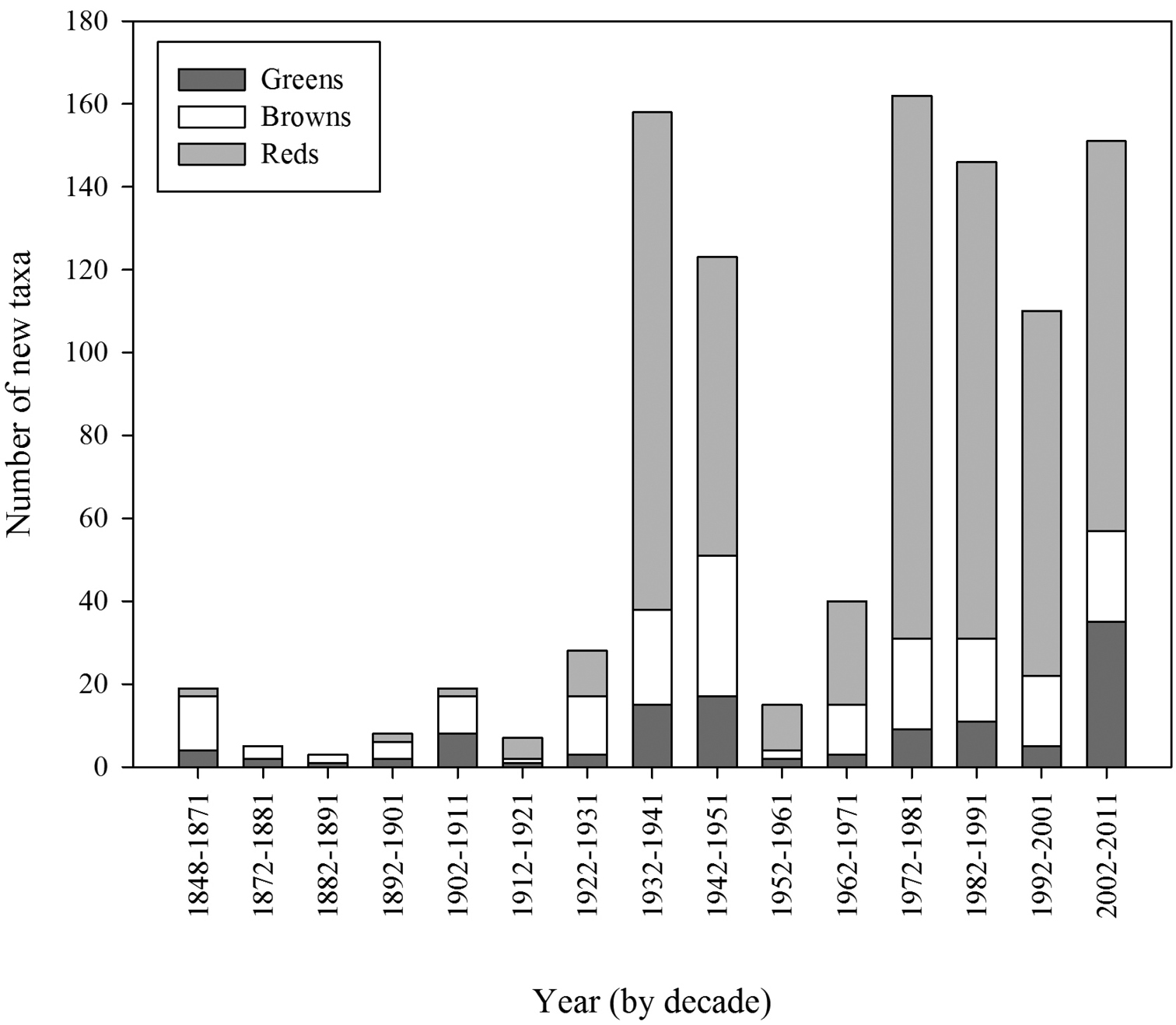
Number of taxa in the collections by decade of collection and division (green, brown, and red algae).
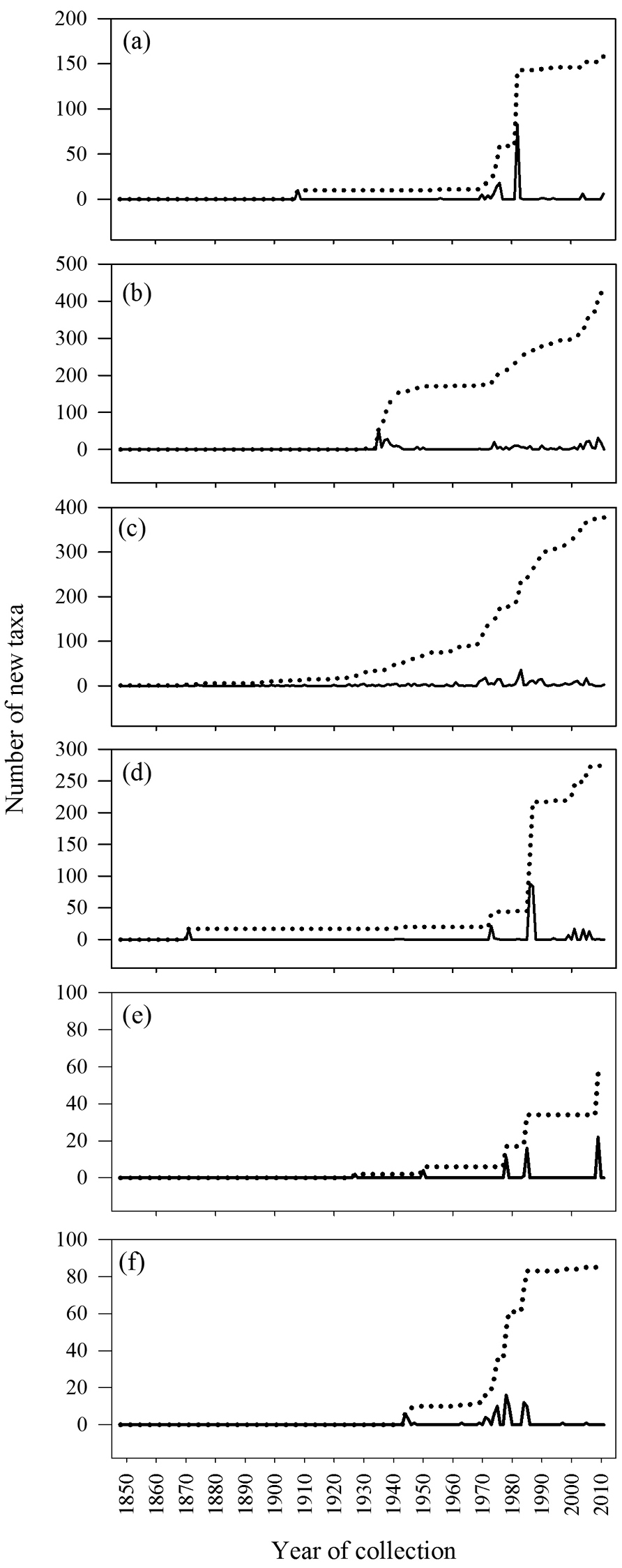
Annual (solid line) and cumulative (dashed line) new taxa from selected regions: a Kermadec Is b NI North c Wairarapa-Cook d Chatham Is e Bounty Is f Campbell I.
Analysis revealed that many entities in the flora are known from very few records (Table 3). Of the 996 taxa in this analysis there are only 210 taxa in the collection for which there are more than 30 records (20 greens, 51 browns, 139 reds). Over the whole collection 17% of the taxon records are known from a single record and 44% from five or fewer records.
Number of taxa and records by class of algae.
| Greens | Browns | Reds | Total | |
|---|---|---|---|---|
| Number of taxa: | 118 | 199 | 679 | 996 |
| Number of taxa known from a single record: | 26 (22%) | 42 (21%) | 97 (14%) | 165 (17%) |
| Number of taxa known from 5 or fewer records | 65 (55%) | 84 (42%) | 294 (43%) | 443 (44%) |
| Number of taxa known from > 30 specimens | 20 (17%) | 51 (25%) | 139 (20%) | 210 (14%) |
| Number of taxa known from >100 records | 3 (3%) | 8 (4%) | 15 (2%) | 26 (3%) |
The data available for each of the 20 regions within New Zealand are summarised in Table 4. This lists the number of taxa, the composition of the flora in each region, the number of records that the data are based on, the composition of the flora by region in terms of the number of classes, orders and families represented, the number of years in which collections were made (of a potential 164 years between 1848-2011), and the seasons in which collecting events occurred. In some regions the collections have been made over an extended period, and span all seasons, whereas it is clear that some other regions are infrequently visited and no collections made in some seasons. Overall the impact of season is relatively modest with the number of collections from the winter months being only two thirds of the total from the summer months. Figure 6 summarises the number of taxa unique to each region by class.
Distribution of macroalgal collections by region.
| Kermadec | 3 Kings | NI North | NI BOP | NI East | NI West | Wairarapa_Cook | SI NW | SI Kaikoura | Chathams | SI SE | SI Westland | SI Fiordland | SI Southern | Stewart | Snares | Bounty | Antipodes | Auckland | Campbell | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Region (Fig. 1) | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 |
| Number of taxa | 159 | 178 | 421 | 314 | 127 | 205 | 381 | 173 | 233 | 275 | 276 | 195 | 271 | 145 | 282 | 111 | 56 | 140 | 101 | 86 |
| Number of greens | 22 | 19 | 61 | 37 | 14 | 21 | 42 | 22 | 24 | 27 | 43 | 19 | 32 | 16 | 31 | 8 | 5 | 17 | 12 | 9 |
| Number of browns | 35 | 36 | 86 | 77 | 34 | 41 | 93 | 32 | 57 | 61 | 60 | 41 | 53 | 31 | 81 | 20 | 10 | 23 | 19 | 16 |
| Number of reds | 102 | 123 | 274 | 200 | 79 | 143 | 246 | 119 | 152 | 187 | 173 | 135 | 186 | 98 | 170 | 83 | 41 | 100 | 70 | 61 |
| Regional taxa as % of total flora | 16.1 | 18.0 | 42.5 | 31.7 | 12.8 | 20.7 | 38.5 | 17.5 | 23.5 | 27.8 | 27.9 | 19.7 | 27.4 | 14.6 | 28.5 | 11.2 | 5.7 | 14.1 | 10.2 | 8.7 |
| Number of unique taxa | 72 | 14 | 35 | 8 | 1 | 6 | 8 | 0 | 2 | 8 | 9 | 0 | 3 | 1 | 11 | 3 | 2 | 5 | 4 | 1 |
| Number of records | 532 | 714 | 2998 | 1372 | 339 | 645 | 3222 | 565 | 659 | 1527 | 1070 | 918 | 1588 | 407 | 1289 | 310 | 147 | 552 | 349 | 219 |
| Number of taxa known from: a single record (%) | 63 (40) | 61 (34) | 104 (25) | 93 (30) | 40 (31) | 77 (38) | 73 (19) | 49 (28) | 97 (42) | 50 (18) | 86 (31) | 33 (17) | 69 (25) | 52 (36) | 61 (22) | 46 (41) | 22 (39) | 52 (37) | 36 (36) | 38 (44) |
| Number of taxa known from 5 or fewer records (%) | 131 (82) | 134 (75) | 254 (60) | 237 (75) | 116 (91) | 177 (86) | 208 (55) | 144 (83) | 207 (89) | 170 (62) | 222 (80) | 143 (73) | 180 (66) | 130 (90) | 195 (69) | 97 (87) | 52 (93) | 105 (75) | 83 (82) | 76 (88) |
| Number of taxa known from > 30 specimens | 0 | 1 | 10 | 4 | 0 | 0 | 14 | 0 | 0 | 0 | 0 | 1 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Collecting events | 92 | 74 | 707 | 420 | 84 | 226 | 957 | 93 | 261 | 196 | 253 | 110 | 272 | 84 | 272 | 59 | 28 | 79 | 84 | 46 |
| Number of years collected | 18 | 13 | 60 | 54 | 23 | 48 | 87 | 22 | 47 | 23 | 56 | 26 | 30 | 28 | 47 | 14 | 6 | 9 | 19 | 16 |
| Springs collected | 7 | 5 | 37 | 30 | 11 | 19 | 53 | 5 | 24 | 4 | 28 | 15 | 10 | 8 | 26 | 6 | 2 | 4 | 4 | 7 |
| Summers collected | 1 | 9 | 40 | 29 | 13 | 22 | 54 | 14 | 19 | 16 | 31 | 17 | 21 | 15 | 31 | 13 | 0 | 2 | 15 | 8 |
| Autumns collected | 9 | 3 | 33 | 32 | 5 | 25 | 55 | 8 | 18 | 7 | 20 | 7 | 10 | 8 | 19 | 1 | 4 | 4 | 3 | 5 |
| Winters collected | 5 | 0 | 26 | 24 | 5 | 22 | 47 | 3 | 21 | 5 | 17 | 6 | 6 | 7 | 14 | 1 | 0 | 0 | 2 | 1 |
| Number of classes | 5 | 6 | 8 | 5 | 4 | 5 | 6 | 5 | 7 | 6 | 6 | 4 | 5 | 5 | 7 | 4 | 5 | 6 | 6 | 5 |
| Number of orders | 25 | 30 | 36 | 30 | 24 | 29 | 31 | 28 | 33 | 32 | 33 | 29 | 30 | 25 | 32 | 26 | 19 | 29 | 27 | 25 |
| Number of families | 50 | 55 | 83 | 73 | 44 | 57 | 75 | 53 | 64 | 72 | 67 | 59 | 62 | 49 | 69 | 44 | 29 | 46 | 48 | 42 |
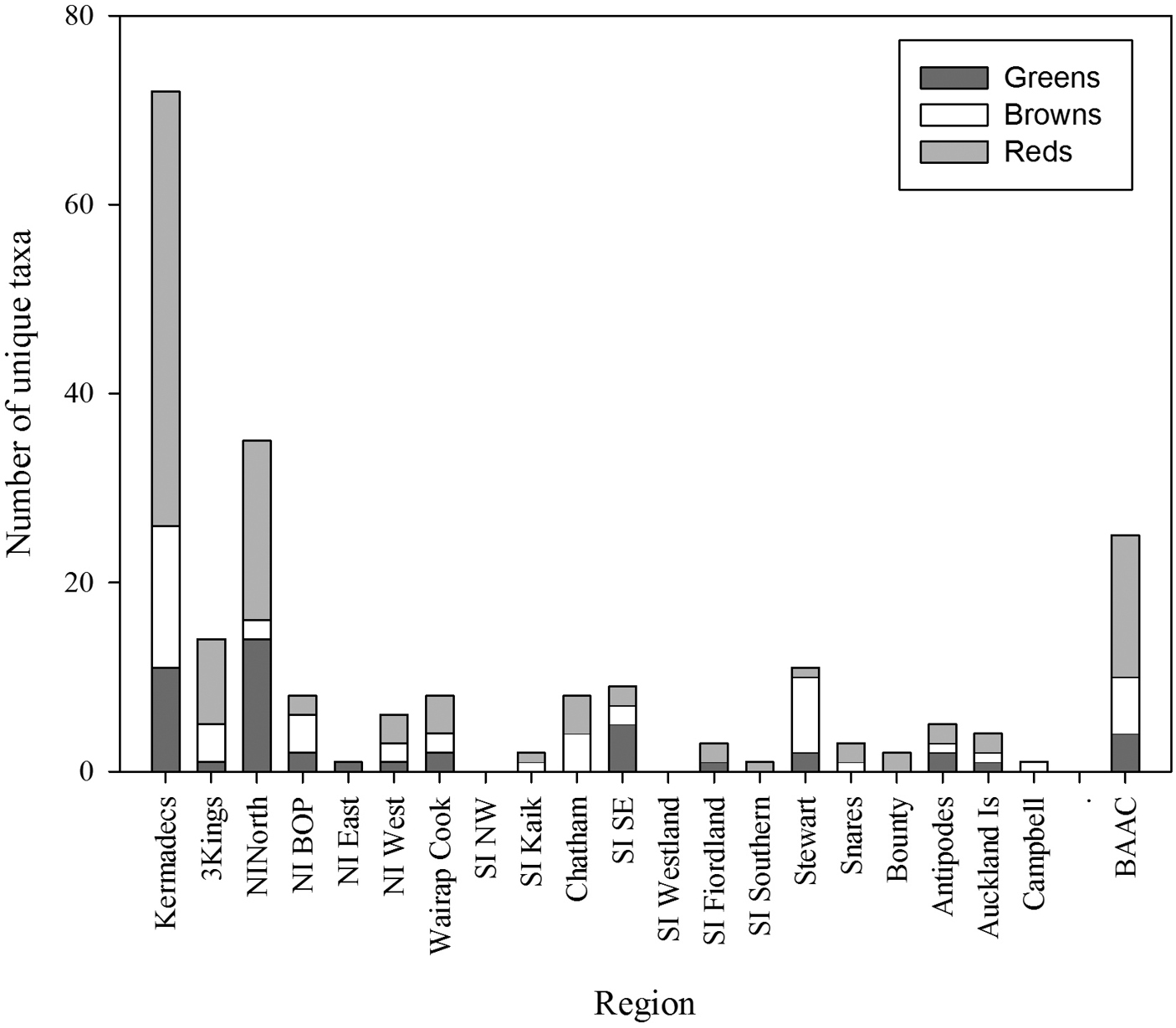
Number of taxa unique to each region by division (BAAC = Bounty, Antipodes, Auckland, Campbell Islands).
These analyses have provided an opportunity to review the state of knowledge of the New Zealand macroalgal flora and to investigate how well the current collections at Te Papa represent the macroalgal flora of New Zealand. The cumulative total of taxa in WELT shows that the flora is not reaching asymptote, suggesting that more discoveries are likely with further investigations of the flora. It is clear that some regions of New Zealand have received greater attention (number of records, collecting events) (Table 4), and as a consequence the flora is better understood in these areas (in particular Wairarapa-Cook, NI North). The number of collections available by region is in large part a consequence of the presence of active research workers, research institutions or programmes, as well as the accessibility of the coastline. The northern and southern island groups - Kermadec and Three Kings Islands, and Snares, Bounty, Antipodes, Auckland, Campbell Islands - are all difficult to reach, subject to inclement weather and are relatively infrequently sampled, i.e. collections made in fewer than 20 years in the 164 years since the first New Zealand collections were lodged in WELT (Table 4).
The knowledge of the flora in different regions has been built up in quite different ways. Macroalgae were first collected from the Kermadec Islands by New Zealand based scientists in 1908, although material had been collected in 1854 and 1874 by expeditions and lodged in European herbaria (
The collection history of the NI North (Fig. 5b) and Wairarapa-Cook (Fig. 5c) are interesting to compare. These are the most diverse and intensely sampled regions. The Bay of Islands is a key collecting area in the NI North and has seen bursts of collecting activities by Tilden and Lindauer in particular. The collecting history of the Wairarapa-Cook region reflects the presence of phycologists and collectors, with a steady growth in the knowledge and representation of the flora over an extended period. The first collections of macroalgae from the Chatham Islands that formed the basis of a published account were made in 1863-64 and then in 1871, with further collections made by a German research expedition in 1897. However it was not until the mid-1980s that thorough and detailed collections were made of the flora (Fig. 5d). In the intervening 80-90 years there were only scattered and infrequent collections made on the islands (summarised in
In terms of regional diversity the northern North Island has the most taxa recorded, contains the highest proportion of the total flora, and has the greatest phylogenetic diversity (as represented by the highest numbers of classes, orders and families present). Although there are almost 3000 specimen records from the region, 25% of the taxa from the area are known from a single record, and 60% from 5 or fewer records. The offshore Three Kings Islands and the southern islands (Snares, Bounty, Antipodes, Auckland and Campbell Islands) are represented by the smallest number of collections resulting from few collecting events. The mainland areas that have received the least collecting effort (fewest collection events) are North Island East, South Island Northwest, and the South Island Southern.
The proportion of the flora that is represented by a very small number of records is salutary, with ca. 44% of the flora known from 5 or fewer records (Table 3). When the data are examined by region (Table 4) the differences in the coverage of collections can be assessed. For phenological studies and comparative investigations it is important to have a number of specimens to evaluate variation and attributes that may be influenced by maturity, seasonality, and/or reproductive status. There are 210 taxa in the collections, identified to species, for which there are more than 30 records: in our view, this number of records provides sufficient individuals for such comparative studies. In terms of the application of NHC collections for understanding the responses of the flora to human-induced environmental changes,
Our analyses have enabled us to test the quality of the data associated with the specimens. The data grooming exercises prior to these analyses have minimised location errors (e.g. latitude and longitude, place names) but opportunity for minor transcription errors still remains. The main issues affecting data quality are the level and standard of identification, which are directly influenced by the current state of the systematic knowledge of the flora. There have been few monographic studies of macroalgal taxa in New Zealand, but recent research across a range of orders has revealed new taxa, and the need for significant taxonomic revisions. These studies have also concluded that understanding the diversity in the flora is still in a discovery phase (e.g.
Although it is questionable whether the Te Papa collections constitute a comprehensive or sufficient baseline with which to evaluate change in the environment or in the flora composition, these herbarium specimens are a very significant source of data both for current biodiversity assessment and planning and also for future applications in biodiversity analysis, conservation and ecology. There are areas within the collection that have been developed from specific research programmes (e.g. collections for the regional floral list series, Bangiales, coralline algae, macroalgae from soft sediment environments, Ulvaceae) where the specimens have been collected in a systematic and targeted way, and in some cases can be associated with other key environmental data. These collections provide a reference baseline if there are opportunities for the regions or habitats to be resampled in the future. The analyses performed here are repeatable if the collection continues to receive the same attention, i.e., expert identification and application of current names, precision of data entry with respect to consistency of place names, and coordinate data
This analysis has identified gaps in the macroalgal collections, both taxonomically and geographically, and also data that can inform future collection development. A number of recent papers reviewing the role of NHCs have stressed the function of museums and herbaria as “part of the essential infrastructure of science” (
This research was funded by NIWA under Coasts and Oceans Research Programmes 2 and 3 (2012/13 SCI). Erika Mackay is thanked for preparing Figure 1.