





(C) 2013 Geoffrey A. Levin. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Levin GA (2013) A synopsis of the New World species of Drypetes section Drypetes (Putranjivaceae) with asymmetrical fruits, including description of a new species. PhytoKeys 29: 75–87. doi: 10.3897/phytokeys.29.6004
A synopsis of the New World species of Drypetes (Putranjivaceae) with asymmetrical drupes is presented. The group consists of three species: Drypetes alba, with two varieties, from the West Indies, Drypetes gentryi from Mexico, and the newly described Drypetes asymmetricarpafrom Costa Rica. The new species can be distinguished from both its relatives by its longer fruiting pedicels. In addition, the new species differs from Drypetes alba by its larger fruits, and from Drypetes gentryi by having shorter staminate pedicels and stigmas borne on styles (rather than sessile). Lectotypes are designated for Drypetes albavar. latifoliaand Drypetes incurva.
Costa Rica, Drypetes, Mexico, Putranjivaceae, West Indies
The genus Drypetes Vahl (Putranjivaceae) contains about 220 species of dioecious trees and shrubs, mostly of the Old World tropics. About 17 known species are found in the Americas, with the greatest diversity in the West Indies. However the Amazonian species are poorly studied and further research undoubtedly will yield many new species. In the classification of
Although no formal groups below the sectional level have been recognized among New World members of section Drypetes, there is a distinctive group of species with strongly asymmetrical drupes. In these species, the young ovary is symmetrical, as in other members of section Drypetes, but as the fruit develops the ovary grows faster on one side than the other, resulting in an oblique fruit apex with the stigma shifted to one side (Fig. 1B). As many as four species commonly have been recognized in this group. Here these are reduced to two species, one with two varieties, and a new species is described. The south Asian species Drypetes gardneri (Thwaites) Pax & K. Hoffm., Drypetes lanceolata (Thwaites) Pax & K. Hoffm., and Drypetes venusta (Wight) Pax & K. Hoffm. have somewhat similar fruits but differ significantly from the American species in foliar and floral characters and probably are not closely related.
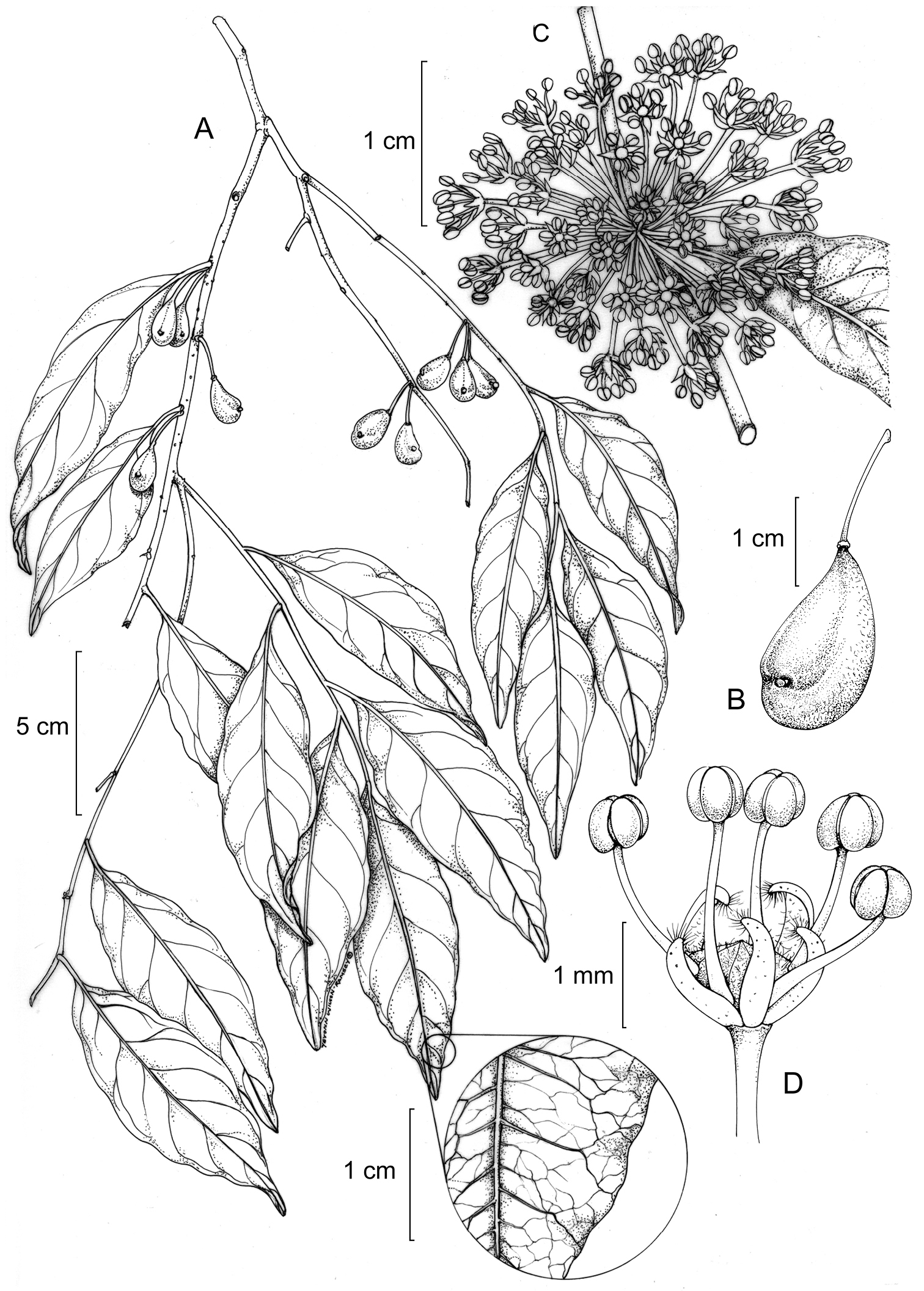
Drypetes asymmetricarpa. A Fruiting branch (detail of venation) B Immature fruit C Staminate inflorescence D Staminate flower. (A–B from Grayum 6614, MO; C–D from Harmon 41, MO).
The study was based on the examination of specimens from the following herbaria: A, ARIZ, BM, CAS, CM, CR, DAV, F, G, G-DC, GH, GOET, HAJB, ILLS, K, LL, MEXU, MICH, MO, NY, P, SD, TEX, U, UC, UCR, and US. All cited specimens were seen by the author unless otherwise indicated. Identifications were made by comparison with the original descriptions and, when available, with the type material or photographs of types. Details of the flowers and fruits were examined under a stereoscopic microscope. All descriptions and data on flowering times, habitats, and distribution are based on the herbarium material examined. Countries in the Selected Specimens sections are listed alphabetically.
| 1 | Fruiting pedicels (8–)10–15 mm long; Costa Rica | 2. Drypetes asymmetricarpa |
| – | Fruiting pedicels 4–10 mm long; Mexico and West Indies | |
| 2 | Staminate pedicels 7–14 mm long; drupes 12–15 mm long; Mexico | 3. Drypetes gentryi |
| – | Staminate pedicels 3–6 mm long; drupes 9–10(–13) mm long; West Indies | [1. Drypetes alba] |
| 3 | Stigmas sessile; Hispaniola, Puerto Rico, U.S. Virgin Islands | 1a. Drypetes alba var. alba |
| – | Stigmas borne on style 1 mm long; Cuba, Jamaica, Lesser Antilles | 1b. Drypetes alba var. latifolia |
[Haiti.] Île de Tortue, s.d., A. Poiteau s.n. (holotype: P; isotype: G-DC).
Haiti, Dominican Republic, Puerto Rico, and U.S. Virgin Islands.
Forests from sea level to 1000 m, primarily on calcareous soils, but in Puerto Rico also on serpentine-derived soils.
Flowering primarily January to April, rarely as early as November. Fruiting February to August.
Least concern. Drypetes alba var. alba is widespread and likely secure in the Dominican Republic, Puerto Rico, and U.S. Virgin Islands. It may be vulnerable or endangered in Haiti due to extensive deforestation there; I have seen no specimens from Haiti collected later than 1929.
HAITI. Southeast of St. Louis du Nord, 3 Apr 1928 (♂ fl), Leonard & Leonard 14268 (CM, K, UC, US).
Dominican Republic. Loma Mala, near arroyo Guayabal, Maimón, 300–500 m, 16 Feb 1974 (fr), Liogier 21293 (F, K, MO).
Puerto Rico. Maricao Afuera, along Maricao River upstream from the fish hatchery, 18°10'11"N, 66°59'10"W, 460–580 m, 11 Jan 1996 (♀ fl), Acevedo-Rodríguez et al. 7724 (K, US).
United States Virgin Islands. Saint Croix: Salt River bluffs, 6 Jul 1896 (♂ fl), Ricksecker 462 (P). Saint John: Coral Bay Quarter, Bordeaux Mtn., east side, 11 Jan 1992 (bud), Acevedo-Rodríguez & Siaca 4710 (MO, US). Saint Thomas: s.d. (fr), Riedlé s.n. (F, P).
Cuba, occ., 1863 (fr), Wright 1927 (lectotype, designated here: GOET003380, photo plants.jstor.org/specimen/goet003380; duplicates: GH pro parte, K pro parte, MO pro parte, NY pro parte).
Cuba, Jamaica, and the Lesser Antilles (Antigua, Guadeloupe, Martinique, Barbados).
Forests on limestone and schist, from sea level to 1100 m.
Flowering October to March. Fruiting December to July.
Least concern. Drypetes alba var. latifolia is widespread and probably secure in Cuba and Jamaica. Its status in the Lesser Antilles is difficult to assess because of a paucity of specimens.
The plants I include in Drypetes alba have been segregated into species or varieties in various ways since the middle of the 19th century (
The only character that seems consistently to differentiate Drypetes alba var. alba from var. latifolia is the presence of a style about 1 mm long in the former and its absence in the latter, the stigma being sessile. This character was first observed by
CUBA. Camagüey: Banao, 300–500 m, Nov 1975 (♂ fl), Alvarez et al. 28778 (HAJB). Guantánamo: San Antonia del Sur, Puriales de Caujeri, Sierra de Purial cerca de Arroyo, 800 m, 30 May 1982 (fr), Bisse et al. 47259 (HAJB). Holguín: Sierra de Nipe, prope Río Piloto, 350 m, 16 Dec 1915 (♂ fl), Ekman 6694 (F, K, NY, U, US). Isla de la Juventud: Caleta Cocodrilos, 8 Mar 1916 (fr), Britton et al. 15305 (CM, F, NY, US). Matanzas: Cienega, Peninsula de Zapata, montes al norte de Sto. Tomás, 19 Apr 1977 (fr), Bisse et al. 34469 (HAJB). Pinar del Río: La Guásima, Rangel, Jan 1950 (♀ fl), Liogier 1261 (GH, US). Sancti Spíritus: Trinidad Mountains, Arroyo Grande, 650–750 m, 11–12 Mar 1910 (fr), Britton &Wilson 5459 (F, NY). Santiago de Cuba: Bayate, 20 Feb 1917 (fr), Ekman 8544 (F, K, LL, NY, U, US).
Jamaica. Trelawny: Boothe district, ca. 3 mi. north of Troy, 1600 m, 14 Mar 1955 (♀ fl, fr), Proctor 9956 (NY, US).
Antigua and Barbuda. Antigua: environs de St. Jean, Dec 1902 (♂ fl), Duss 80 (NY).
Guadeloupe. Marie-Galante, bois de Folle-Anse, 1896 (fr), Duss 3628 (F, MO pro parte, NY, US pro parte).
Martinique. Morne Saint-Martin, pied de la montagne Pelée, 1878 (♂ fl), Duss 50 (NY).
urn:lsid:ipni.org:names:77134216-1
http://species-id.net/wiki/Drypetes_asymmetricarpa
Figure 1Differs from the other New World Drypetes species withasymmetrical fruits by its longer fruiting pedicels [(8–)10–15 mm vs. 4–10 mm]; also differs from Drypetes alba by its larger fruits [12–18 mm vs. 9–10(–13) mm], and from Drypetes gentryi by having shorter staminate pedicels (5–8 mm vs. 7–14 mm) and stigmas borne on styles (vs. being sessile).
Costa Rica. Puntarenas: Cove at NE base of peninsula, Punta Quepos (3 km S of Puerto Quepos), 9°24'N, 84°10'W, 0 m, 8 Mar 1986 (fr), M. H. Grayum 6614 (holotype: MO, isotypes: CR n.v., F).
Trees 6–20 m, to 35 cm dbh; bark with longitudinal fissures; branches brown when young, becoming gray, glabrous or sparsely minutely puberulent with spreading hairs. Leaves: stipules 0.5 × 0.5 mm, deltate, puberulent; petiole 3–10 × 0.3–1 mm, glabrous; blade elliptic to lanceolate, straight or somewhat curved, 4–12 × 1.5–4.5 cm, base asymmetrical, acute, margins entire or minutely crenulate-serrulate, often undulate, apex attenuate, surfaces glabrous, 2° veins 6–9/side. Inflorescences axillary fascicles; staminate 25–40-flowered, bracts 0.5 mm, puberulent, pedicels 5–8 × 0.2 mm, glabrous; pistillate (known only in fruit) 2–6-flowered, bracts 0.25 × 0.25 mm, deltate, puberulent, pedicels (8–)10–15 × 0.4–0.8 mm, glabrous. Staminate flowers: sepals 5–6, narrowly triangular to narrowly lanceolate, 1 × 0.3 mm, spreading and slightly incurved at apex, apex acute, margins ciliate, abaxial surface glabrous except puberulent at apex, adaxial surface densely to sparsely puberulent; stamens 5(–6), irregularly alternate and opposite sepals, filaments 1.5–2 mm × 0.1 mm, glabrous, anthers 0.4–0.5 × 0.4–0.5 mm, glabrous, latrorse; disc lobed between stamens, densely puberulent. Pistillate flowers unknown, but remnant sepals (below fruits) ovate-elliptic, 1.5 × 0.6 mm, apex acute and slightly incurved, abaxial surface glabrous, adaxial surface densely puberulent; disc annular, densely puberulent; ovary unknown; style becoming subapical during fruit development, 0.5 mm; stigma subreniform, 0.5 × 1 mm, glabrous. Drupes (immature) green, 1-carpellate, ovoid-globose, 12–18 × 7–10 × 6–8 mm, apex strongly asymmetrical, sparsely to densely puberulent with very short hairs (0.1 mm). Seed 1.
The specific epithet refers to the strongly asymmetrical drupes, which are unique among Central American Drypetes.
Known only from Costa Rica, where it is found from the north central part of the country to the central west coast. It may also be expected in extreme southern Nicaragua.
Forests at elevations from sea level to 750 m.
Flowering January (possibly longer, but only a single flowering specimen known). Fruiting March to June (possibly longer as only immature fruits are known).
Probably of Least Concern. The range of Drypetes asymmetricarpa spans at least 200 km. Although its habitat is highly fragmented, the species is found in Manuel Antonio National Park and near both Guanacaste and Rincón de la Vieja national parks.
Drypetes asymmetricarpa was listed by
All the pistillate specimens studied have immature fruits, so their full size and color at maturity are unknown. The label on Hammel & Trainer 17046, with fruits 9–11 × 7–8 × 6–7 mm, says “fruits ca. 1/3 full size, ” but based on my experience with other species I suspect this overestimates the mature size.
COSTA RICA. Alajuela: Cantón de Upala, Distrito Dos Ríos, 7.5 km NE of town, between La Jabalina and the Río Cucaracho, 10°56'N, 85°19'W, 325 m, 4 Apr 1988 (fr), Herrera 1693 (F, ILLS, MO); Puntarenas: Cantón de Puntarenas, Distrito Monteverde, San Luis, finca de Chepe Rojas, al oeste del pueblo, 10°16'N, 84°50'W, 750 m, 24 Jun 1988 (fr), Bello et al. 35 (F, ILLS, MO), Bello et al. 58 (F, ILLS, MO); Monteverde area from Santa Elena to San Luis, 10°16'N, 84°50'W, 700 m, 16 Jun 1988 (fr), Hammel & Trainer 17046 (F, ILLS, MO); Parque Nacional Manuel Antonio, Playa Espadilla Sur. 9°24'N, 84°10'W, 1–100 m. 2 Jan 1990 (♂ fl), Harmon 41 (MO).
Mexico. Sinaloa: Capadero, Sierra Tacuichamona, rocky canyon under basaltic rim, 3500 ft., 13 Feb 1940 (fr), H. S. Gentry 5597 (holotype: NY, isotypes: ARIZ, MICH, MO).
Western Mexico, in the Sierra Madre Occidental from near 27° N in Chihuahua and Sonora to about 19° N in Colima.
Tropical deciduous forests at about 100–1100m.
Flowering December–February. Fruiting December–June.
Least Concern. Drypetes gentryi is widespread in the lower elevations of the Sierra Madre Occidental.
When
Trees 8–25 m, often with multiple trunks from near base, to 20–100 cm dbh; bark scaled and with longitudinal fissures; branches brown when young, becoming gray, minutely puberulent with spreading hairs, becoming glabrous. Leaves: stipules 0.5–0.6 × 0.7–1 mm, deltate, puberulent; petiole 6–12 × 0.7–1 mm, puberulent with spreading hairs or glabrous; blade elliptic to lanceolate, straight or somewhat curved, 4–15 × 1.5–4 cm, base asymmetrical, acute to narrowly obtuse, margins subentire to crenulate-serrulate, often undulate, apex attenuate, surfaces glabrous or very sparsely pubescent with appressed hairs especially near base, 2° veins 6–9/side. Inflorescences axillary fascicles; staminate 20–40-flowered, bracts 0.5 × 0.5 mm, deltate, puberulent, pedicels 7–14 × 0.2 mm, glabrous; pistillate 1–6-flowered, bracts 0.5 x0.5 mm, deltate, puberulent, pedicels 3–10 × 0.4–0.5 mm, puberulent when young, becoming glabrous. Staminate flowers: sepals 5(–6), linear to narrowly triangular, 1.2 × 0.4 mm, spreading and slightly incurved at apex, apex bluntly acute, margins ciliate, abaxial surface glabrous except puberulent at apex, adaxial surface puberulent; stamens 5(–6), mostly opposite sepals, filaments 1.6–2.2 mm × 0.1 mm, glabrous, anthers 0.8–1 × 0.5–0.6 mm, glabrous, latrorse; disc lobed between stamens, densely puberulent. Pistillate flowers: sepals 5, narrowly triangular to linear, 1–1.2 × 0.3–0.4 mm, spreading, entire, apex bluntly acute, abaxial surface glabrous to sparsely puberulent but densely puberulent at apex, adaxial surface densely puberulent; disc annular, densely puberulent; ovary densely puberulent; style absent; stigma apical at anthesis, becoming subapical during fruit development, subreniform, 0.8 × 1.2 mm, glabrous. Drupes (immature) green, 1-carpellate, obovoid, 12–15 × 7–9 × 6–8 mm, apex strongly asymmetrical, densely puberulent with very short hairs (0.1 mm). Seed 1.
The mature fruits are described as white (Bye 6066) or yellow (Bye et al. 12847), with the mesocarp juicy and both sweet and astringent (Bye et al. 12847). Spanish vernacular names include cortopico (Gentry 5597), palo masiso (Bye 9707), and tempisque (Bye 3401, Bye et al. 12847); in Tarahumara it is called bapible (Bye 3401) or kafe (Bye et al. 12847), and in Guarijio joyarí (Felger et al. 94-56).
MEXICO. Chihuahua: Mpio. Batopilas, north side of Barranca de Batopilas, along arroyo Samachique between Rio Batopilas and Tarahuamara village of Wimivo, 27°09'N, 107°38'W, 900–1000 m, 30 May 1980 (fr), Bye 9707 (ARIZ, DAV, F, GH, ILLS, MEXU, MICH, MO, NY, SD, TEX, UCR, US); Colima: canyon near Rio Marabasco (Cihuatlan) bridge on road to Chacala, north of Santiago, 19°17'N, 104°19'W, 200–250 m, 21 Jan 1988 (fr), Levin & Dice 1975 (MO, SD); Jalisco: canyon east of Highway 200 ca. 2 km east-southeast of Boca de Tomatlan, at bridge, 20°03'N, 105°18'W, 100–200 m, 25 Jan 1988 (♂), Levin & Dice 2001 (MO, SD); Sonora: Arroyo Gochico ca. 8 km E of San Bernardo, 27°02'04"N, 108°04'07"W, 300 m, 31 Jan 1988 (♀ fl, fr) Levin et al. 2015 (MO, SD).
I thank Jochen Heinrichs for providing high-resolution scans of specimens at GOET, and the staff of the other herbaria listed for providing access to the specimens in their care. Lynn Gillespie and three anonymous reviewers provided helpful comments on the manuscript. Figure 1 was prepared by Yevonn Wilson-Ramsey.
Abbott, W.L. 2571, 2932 (alba var. alba)
Acevedo-Rodríguez, P. 4710, 4781, 7724 (alba var. alba)
Alvarez, A. 28778, 45576 (alba var. latifolia)
Areces, A. 25812 (alba var. latifolia)
Bello, E. 35, 58 (asymmetricarpa)
Bisse, J. 16899, 22625, 26478, 34469, 46192, 47203, 47259, 53290 (alba var. latifolia)
Britton, E.G. 5154 (alba var. alba)
Britton, N.L. 2457, 2748, 4128, 4208 (alba var. alba); 5459 (alba var. latifolia); 8104, 9676 (alba var. alba); 15305 (alba var. latifolia)
Bye, R.A. 3401, 3557, 3558, 3958, 6066, 9707, 12847 (gentryi)
Duke, J.A. 7350 (alba var. alba)
Duss, A. 50, 80 (alba var. latifolia); 3628 (alba var. latifolia and Drypetes glauca Vahl); 4142 (alba var. latifolia)
Eggers, H.F.A. 2612 (alba var. alba)
Ekman, E.L. 4146 (alba var. latifolia); 4179 (alba var. alba); 4543, 4568, 4838, 5006 (alba var. latifolia); 5528 (alba var. alba); 6694, 8544, 9764, 9913 (alba var. latifolia); 12160 (alba var. alba); 12462, 18213 (alba var. latifolia); H.3512, H.3870, H.4178, H.4179, H.5134, H.5528, H.6061, H.9614, H.12160 (alba var. alba)
Felger, R.S. 94-56, 97-118 (gentryi)
Gentry, A.H. 50716 (alba var. alba)
Gentry, H.S. 3618, 5597 (gentryi)
Grayum, M.H. 6614 (asymmetricarpa)
Haenke, T. 1605 (gentryi)
Hammel, B.E. 17046 (asymmetricarpa)
Harmon, W.E. (asymmetricarpa)
Harris, W. 7098 (alba var. latifolia)
Herrera, G. 1693 (asymmetricarpa)
Holdridge, L.R. 149 (alba var. alba)
Jack, J.G. 7809 (alba var. latifolia)
Jiménez Almonte, J. de J. 1981, 4360, 5058 (alba var. alba)
León, Bro. 6081, 11800 (alba var. latifolia)
Leonard, E.C. 11589, 14257, 14268, 15686 (alba var. alba)
Lesueur, D.H. 1419 (gentryi)
Levin, G.A. 1973, 1975, 2001, 2014, 2015 (gentryi)
Liogier, A.H. 1017, 1261, 1275, 2997, 5719 (alba var. latifolia); 10466, 10714, 10779, 14325, 15661, 20858, 21293, 21600, 22628, 24490, 24537, 25846, 26463, 26835 (alba var. alba)
Little, E.L., Jr. 13297, 13366 (alba var. alba)
Luna, A. 15 (alba var. latifolia)
McVaugh, R. 1647, 25404 (gentryi)
Pérez J., L.A. 413 (gentryi)
Proctor, G.R. 9956 (alba var. latifolia)
Ricksecker, A.E. 462 (alba var. alba)
Sagra, R. de la 607 (alba var. latifolia)
Sanders, A.C. 1096 (gentryi)
Schiffino, J. 15 (alba var. alba)
Shafer, J.A. 3693, 4403 (alba var. latifolia)
Sintenis, P.E.E. 1502 (alba var. alba)
Stehlé, H. 416 (alba var. latifolia)
Valeur, E.J. 837 (alba var. alba)
Van Devender, T.R. 93-1381 (gentryi)
Wilson, P. 9287 (alba var. latifolia)
Wright, C. 498, 501 (alba var. alba and Drypetes lateriflora (Sw.) Krug & Urb.); 593 (alba var. latifolia and D. lateriflora); 1112 (alba var. latifolia); 1927 (alba var. latifolia and D. lateriflora); 1928, 1929, 1929, 593b (alba var. latifolia)
Zanoni, T.A. 36401 (alba var. alba)