





(C) 2013 George T. Plunkett. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Plunkett GT, Wilson KL, Bruhl JJ (2013) Sedges in the mist: A new species of Lepidosperma (Cyperaceae, Schoeneae) from the mountains of Tasmania. PhytoKeys 28: 19–59. doi: 10.3897/phytokeys.28.5592
The status of a putative new species of Lepidosperma from the mountains of south-western Tasmania, Australia, was investigated. Phenetic analysis (Flexible UPGMA Agglomerative Hierarchical Fusion and semi-strong hybrid multidimensional scaling) was conducted on a database derived from morphological and anatomical characters scored from herbarium material, culm anatomy slides and scanning electron micrographs of fruit. The results of the analysis support the recognition of a new species, here described as Lepidosperma monticola G.T.Plunkett & J.J.Bruhl. The distribution, habitat and conservation status are discussed.
Lepidosperma inops, Lepidosperma monticola, species limits, phenetics, new species
Lepidosperma Labill. (Cyperaceae) is a genus of c. 75 described species (
The present study was initiated following the inspection by GTP of several specimens that had been identified as Lepidosperma inops F.Muell. ex Rodway. Eight of the specimens, collected in south-western Tasmania, appeared morphologically and ecologically distinct from the rest of the specimens, which were collected in the east of the state and attributable to Lepidosperma inops sens. strict. We intuitively segregated them as a putative new species, referred to as Lepidosperma sp. Eldon Bluff (A.M. Buchanan 9981). Subsequent plant collections in Tasmania and a visit to the Tasmanian Herbarium (HO) further strengthened our initial impression.
The aims of the study were (1) to determine the status and limits of this putative new entity and (2) to investigate its resemblance to morphologically similar named species of Lepidosperma.
A total of 25 specimens were used for the phenetic analysis (Table 1). Lepidosperma curtisiae K.L.Wilson & D.I. Morris, Lepidosperma inops and Lepidosperma tortuosum F. Muell. were sampled on the basis of morphological similarity to Lepidosperma sp. Eldon Bluff. Specimens recently collected in Tasmania and existing specimens from the N.C.W. Beadle Herbarium (NE) were used in the analysis. Specimens on loan from the National Herbarium of Victoria (MEL) and HO for a previous study (
Specimens of Lepidosperma used in phenetic analysis. First three letters of operational taxonomic unit (OTU) code: cur = Lepidosperma curtisiae; eld = Lepidosperma sp. Eldon Bluff; ino = Lepidosperma inops; tor = Lepidosperma tortuosum. Localities are in Tasmania unless another State is specified.
| OTU Code | Voucher |
|---|---|
| curBag01 | Above Bagdad Rivulet, Pontville, A.J. North s.n., 8 October 1996, HO 322622 (HO) |
| curCon01 | Conara, Midlands, J.J. Bruhl 3005 (NE) |
| curMac01 | Macquarie River, 0.5 km SW of Long Marsh Dam, A. Moscal 8040 (HO) |
| curRos01 | Roses Creek Road, Grampians, Victoria, A.C. Beauglehole 25084 (MEL) |
| eldCou01 | Mt Counsel, South West, S.J. Jarman s.n., 12 March 1986, HO 411078 (HO) |
| eldEld01 | Below Eldon Bluff, Central Highlands, A.M. Buchanan 9981 (HO) |
| eldFie01 | Mt Field National Park, Tarn Shelf, G.T. Plunkett 99 (NE) |
| eldFie02 | Mt Field National Park, Tarn Shelf, G.T. Plunkett 100 (NE) |
| eldMcC01 | Mt McCall, West Coast, S.J. Jarman s.n., HO 411443 (HO) |
| eldPro01 | Mt Propsting, South West, M.J. Brown 1365 (HO) |
| eldSpr01 | Mt Sprent, South West, G.T. Plunkett 94a (NE) |
| eldSpr02 | Mt Sprent, South West, G.T. Plunkett 96 (NE) |
| eldSpr03 | Mt Sprent, South West, G.T. Plunkett 98 (NE) |
| inoBre01 | Mills Marsh, Break-O-Day, East Coast, A. Moscal 6182 (NE) |
| inoGra01 | Grasstree Hill, East Coast, A. Moscal 18031 (NE) |
| inoLen01 | Lenah Valley, Midlands, D.A. Ratkowsky 1083 (NE) |
| inoUni01 | University of Tasmania, G.T. Plunkett 112 (NE) |
| inoUni02 | University of Tasmania, G.T. Plunkett 115 (NE) |
| torBaro02 | Barokee Swamp, Cathedral Rock National Park, Northern Tablelands, NSW, J. Hodgon 415 (NE) |
| torBarT01 | Barrington Tops National Park, Northern Tablelands, NSW, J.R. Hosking 3041 (NE) |
| torBla01 | 2 km W of Blackheath, Central Tablelands, NSW, V. Klaphake s.n., 1 January 1998, NE 808939 (NE) |
| torBoo01 | Boonoo Boonoo State Forest, Northern Tablelands, NSW, J.J. Bruhl 2453 (NE) |
| torDem01 | Demon Nature Reserve, Northern Tablelands, NSW, J.T. Hunter 5068 (NE) |
| torMul01 | Surveyors Creek Fire Trail, from Mulligans Drive end, Gibraltar Range National Park, Northern Tablelands, NSW, J. Hodgon 391c (NE) |
| torWer01 | Werrikimbe National Park, Northern Tablelands, NSW, S.J. Griffith s.n., 20 April 1993, NE 66428 (NE) |
Hand-cut cross-sections of culms were made in 50% ethanol using half a double edged razor blade from pickled (70% ethanol) or rehydrated herbarium material. Sections were stained in 1% safranin in 50% ethanol for c. 15 min then washed in increasing concentrations of ethanol before staining in 0.5% fast green in 95% ethanol for three to seven minutes depending on size. Sections were then transferred to increasing concentrations of histolene before mounting on glass slides with Eukitt mountant.
Photosynthetic pathway was inferred from anatomy using the ‘maximum cell distance count’ criterion as applied to Cyperaceae (
Where available, a single representative fruit in good condition was removed from each operational taxonomic unit (OTU). Additional fruit removed from non-OTU specimens were also used. Fruit were mounted onto an aluminium stub with double-sided carbon tabs and gold-coated using a JEOL MP-19020NCTR NeoCoater (JEOL Ltd., Tokyo). Images were captured with a JEOL JCM-5000 benchtop SEM (JEOL Ltd., Tokyo) at 10 kV. In total three fruit were imaged from Lepidosperma curtisiae, four from Lepidosperma inops, seven from Lepidosperma sp. Eldon Bluff and two from Lepidosperma tortuosum.
A character list was constructed primarily from that used by Hodgon (2001) and augmented by
Morphological characters were scored from herbarium specimens with the aid of a Leica M8 stereomicroscope. Morphological dimensions were measured once per OTU, consistently using a steel rule, electronic callipers or the microscope eyepiece graticule for each character. Fruit and perianth characters were assessed under the stereomicroscope and from SEM images. Anatomical characters were scored from permanent double-stained slides viewed under a Zeiss Axiolab compound microscope.
Data were organised using DELTA ver. 1.04 (
Our species concept is consistent with that of
In the phenetic analyses, our criteria for accepting the putative species as a distinct entity were: (1) the OTUs representing Lepidosperma sp. Eldon Bluff should form discrete groups distinct from all other groups of OTUs in the analyses and (2) the OTUs within these groups should show an amount of dissimilarity comparable to that of the known species included in the analyses.
The phenogram produced from the cluster analysis (Figure 1) shows distinct clusters of OTUs that match the three described and one putative species. The levels of dissimilarity within these four groups are comparable. Groups A and B in the phenogram contain individuals of Lepidosperma inops and Lepidosperma sp. Eldon Bluff, respectively. These groups are most similar to one another, as expected, though clearly distinct. The Lepidosperma inops group shows the lowest dissimilarity among individuals, possibly because of the lower number of OTUs used for that species. Group C contains individuals of Lepidosperma curtisiae and shows low dissimilarity among individuals, again possibly related to the low number of OTUs used. Group D contains individuals of Lepidosperma tortuosum and displays the highest dissimilarity among individuals, reflecting the morphological variation sampled in this species.
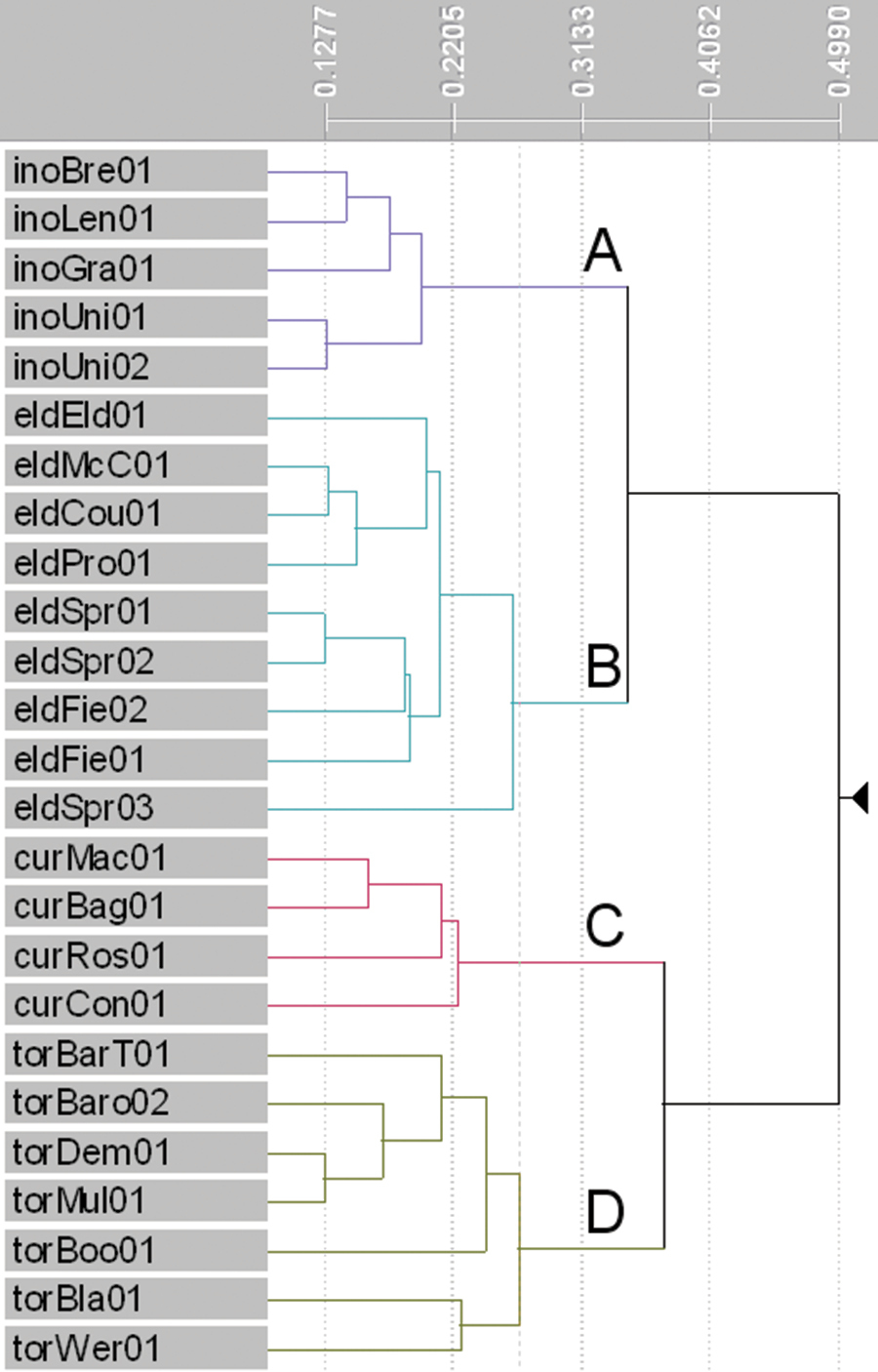
Phenogram produced from cluster analysis of three described and one putative species of Lepidosperma (β = -0.1). Major groups: A Lepidosperma inops (purple) B Lepidosperma sp. Eldon Bluff (turquoise) C Lepidosperma curtisiae (pink) D Lepidosperma tortuosum (green-grey). See Table 1 for operational taxonomic unit codes, Appendix 1 for characters and Appendix 2 for data.
The ordination plot produced from SSH MDS (stress value 0.1535; Figure 2) recovered the same four groups as the phenogram. Individuals of Lepidosperma curtisiae and Lepidosperma inops respectively form the two most closely spaced groups, with individuals of Lepidosperma tortuosum forming the most distantly spaced group. Individuals of Lepidosperma sp. Eldon Bluff form a distinct group that is close to the group containing Lepidosperma inops.
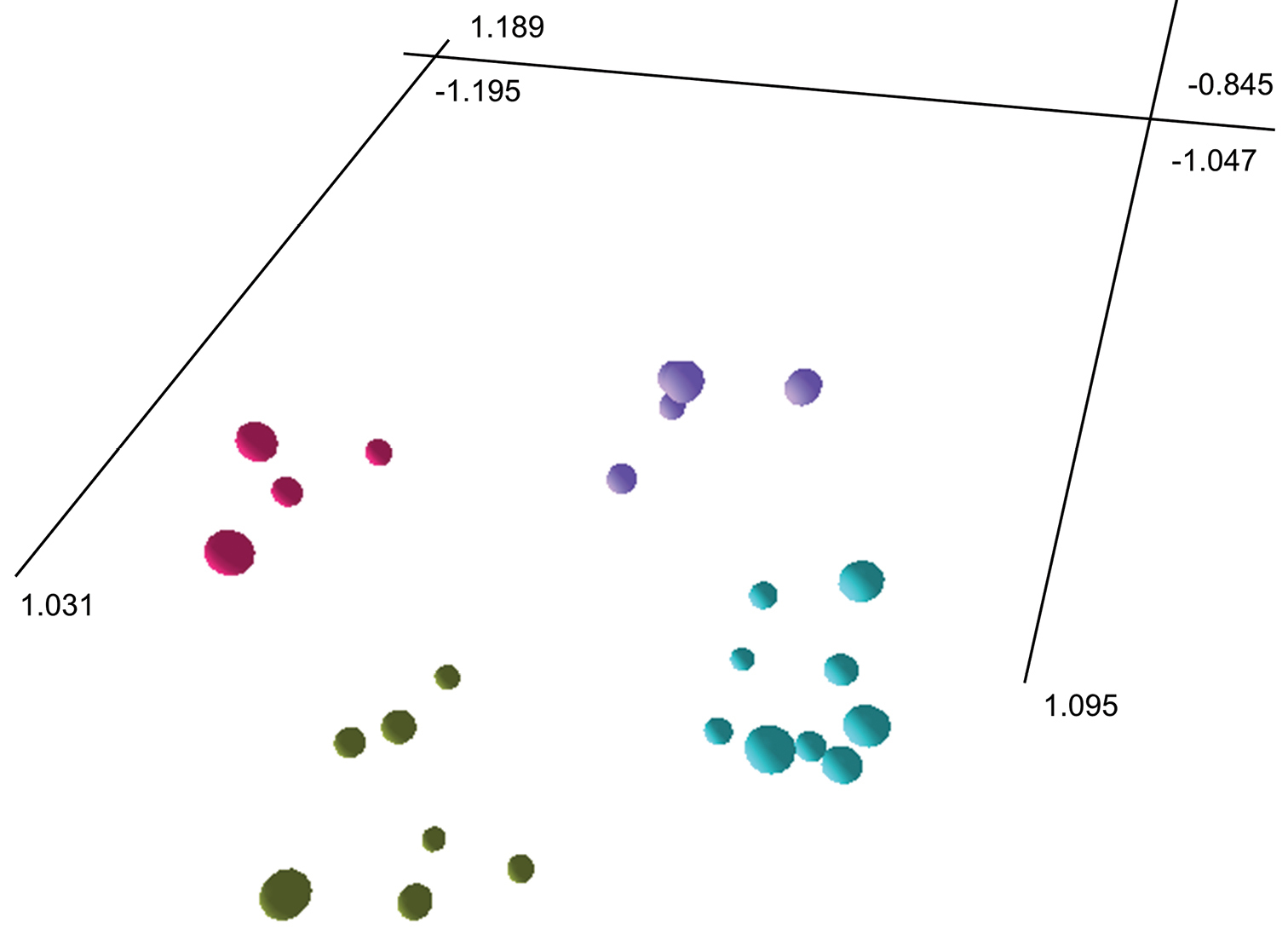
Three-dimensional ordination from semi-strong hybrid multidimenstional scaling of three described and one putative species of Lepidosperma. Lepidosperma curtisiae (top left; pink); Lepidosperma inops (top right; purple); Lepidosperma sp. Eldon Bluff (bottom right; turquoise); Lepidosperma tortuosum (bottom left; green-grey). Ordination oriented to highlight separation of groups of operational taxonomic units. See Appendix 1 for characters and Appendix 2 for data.
The PCC values for the ordination analysis (Table 2) indicate that the most important characters were mostly vegetative and inflorescence morphology characters, with one fruit character and three anatomical characters.
Principal component correlation of characters and ordination vectors from semi-strong hybrid multidimensional scaling ordination of three described and one putative species of Lepidosperma. Characters with a maximum correlation (r2) higher than 0.7 are included. See Appendix 1 for characters and Appendix 2 for data.
| No. | Character | X | Y | Z | r2 |
|---|---|---|---|---|---|
| 128 | Fruit style cap indumentum | -0.718 | 0.3 | 0.628 | 0.845 |
| 9 | Culm margin hairs: orientation | 0.645 | -0.12 | -0.755 | 0.834 |
| 38 | Leaf lamina margin hairs: orientation | 0.407 | -0.667 | -0.624 | 0.767 |
| 2 | Culm length | 0.129 | 0.18 | -0.975 | 0.765 |
| 17 | Angle of ramet | -0.643 | 0.34 | 0.686 | 0.74 |
| 145 | Culm pith: homogeneous or heterogeneous | 0.086 | 0.007 | 0.996 | 0.734 |
| 72 | Rachis: whether reflexed | 0.058 | -0.243 | 0.968 | 0.725 |
| 62 | Involucral bract ligule: emarginate | -0.11 | -0.064 | -0.992 | 0.721 |
| 153 | Culm sub-epidermal fibres: depth | 0.854 | -0.4 | -0.334 | 0.709 |
| 58 | Involucral bract sheath: indumentum | -0.208 | -0.971 | 0.114 | 0.707 |
| 136 | Culm vascular bundles: number of size classes | -0.146 | -0.382 | -0.913 | 0.701 |
The images obtained with SEM demonstrated clear differences between fruits of Lepidosperma sp. Eldon Bluff and its closest neighbour Lepidosperma inops (Figure 3). The fruit of Lepidosperma sp. Eldon Bluff is minutely but distinctly colliculate at its distal end (Figure 3D), while the fruit of Lepidosperma inops is smooth distally (Figure 3B). The size and shape of the fruit and the size of the perianth varies between the two specimens shown here (Figure 3A, C), but no consistent differences were found in perianth size and morphology between the two taxa. Based on our observation of fruit from other species of Lepidosperma, the wrinkled fruit of Lepidosperma inops shown here, although well-developed, is slightly immature and the wrinkling is not diagnostic (old fruits on the same collection in NSW were too fragile to image but still intact enough to see that the main body is only very faintly wrinkled). More mature fruits of Lepidosperma inops suitable to image were not available. Fruits of Lepidosperma curtisiae and Lepidosperma tortuosum are also smooth distally (not shown here). Observations of additional fruit under the stereomicroscope were consistent with the SEM images.
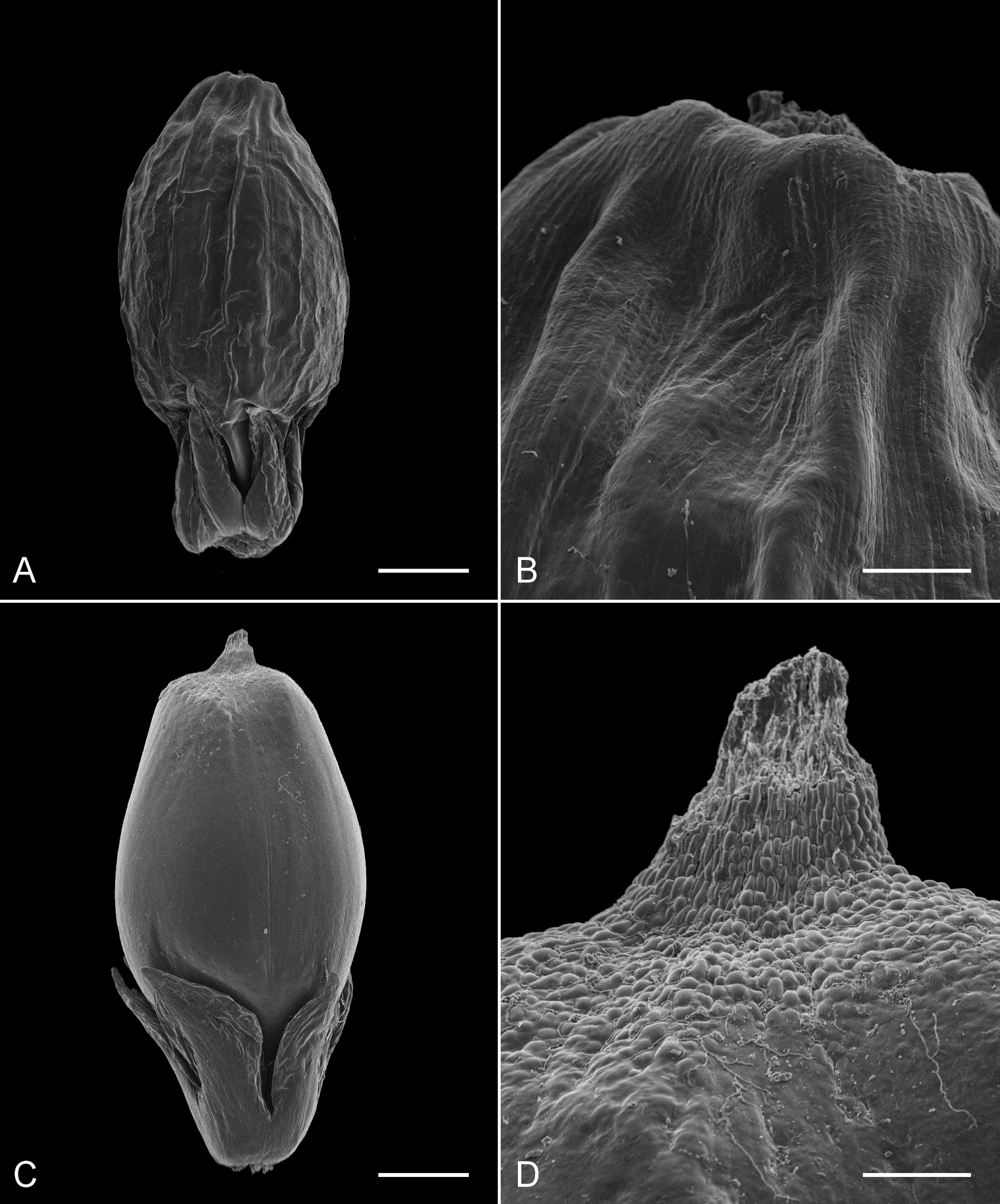
Scanning electron micrographs of fruit with perianth (A, C) and fruit apex (B, D). A, B Lepidosperma inops (A. Moscal 18031) C, D Lepidosperma sp. Eldon Bluff (G.T. Plunkett 100). Scale bars = 0.5 mm (A, C), 0.1 mm (B, D).
The results of the phenetic analysis provide strong support for the acceptance of Lepidosperma sp. Eldon Bluff as a distinct entity. In both the phenogram (Figure 1) and ordination (Figure 2) a discrete group is formed by OTUs representing Lepidosperma sp. Eldon Bluff. This group shows a level of dissimilarity that is comparable to those of the known species of Lepidosperma included in the analysis. The SEM images also show that Lepidosperma sp. Eldon Bluff has distinctive fruit (Figure 3). This evidence provides strong support for the recognition of Lepidosperma sp. Eldon Bluff as a separate species from Lepidosperma inops. The ecological and geographic isolation of these two entities adds further support to this inference (Figure 4; Table 3). Lepidosperma sp. Eldon Bluff is described below as Lepidosperma monticola sp. nov.

Comparison of habitat and habit of Lepidosperma inops and Lepidosperma monticola. A, B Lepidosperma inops (G.T. Plunkett 112) at University of Tasmania campus, Hobart C, D Lepidosperma monticola (G.T. Plunkett 99, J.J. Bruhl & C.J. Prychid) at the type locality, Tarn Shelf, Mt Field National Park. Scale bars = 1 cm. Arrows indicate plants of Lepidosperma inops (A) and Lepidosperma monticola (C).
Selected morphological and ecological attributes separating Lepidosperma monticola and Lepidosperma inops. Ranges presented are absolute.
| Lepidosperma inops | Lepidosperma monticola sp. nov. | |
|---|---|---|
| Morphological character | ||
| Culm length to leaf length ratio | 0.27–0.37 | 0.53–0.7 |
| Involucral bract length | 27–59 mm | 9–25 mm |
| Fruit – distal surface | smooth | minutely colliculate |
| Angle of ramet (i.e. between outermost leaves) | 9–15° | 20–27° |
| Ecology | ||
| Habitat | Grassy woodland or sclerophyll forest | Alpine heath, herbfield, occasionally subalpine woodland |
| Altitude | 20–500 m | 700–1170 m |
^ Measurements from five specimens of Lepidosperma inops and nine of Lepidosperma monticola, fruit observations from four and seven specimens respectively.
† The vegetation at Eldon Bluff for A.M. Buchanan 9981 (HO) at 1080 m is described as “Open Eucalyptus coccifera forest on dolerite talus.”
Lepidosperma monticola is distinguished from Lepidosperma inops in having a minutely colliculate fruit apex, its longest culms being greater than half the length of the leaves, and the angle between the outermost leaves of the ramets being 20° or greater.
AUSTRALIA: Tasmania: Mt Field National Park, Tarn Shelf, c. 100 m N of Dobson Hut, 26 April 2012, G.T. Plunkett 99, J.J. Bruhl & C.J. Prychid; holotype: HO; isotype: AD, BOL, BRI, BRIP, CANB, CHR, GENT, K, MEL, MO, NE, NSW, NY, P, PERTH, PRE. (Figure 5).

Isotype (G.T. Plunkett 99 et al., NE) of Lepidosperma monticola G.T.Plunkett & J.J.Bruhl, sp. nov.
Short-rhizomatous perennial, forming dense clumps. Leaves markedly distichous, up to 8 cm long, innovations forming flat fans (ramets) with angle of 20–27° between outer leaves; lamina well-developed, isobilateral, shallowly biconvex, ± rigid, striate, to 65 mm long, 1–1.5 mm wide, margins glabrous or scabrous with prickle hairs antrorse; sheaths 10–25 mm long, pale yellow-brown to mid-brown, often with tinge of red-pink, not resinous, margin glabrous; ligule subulate to acute, glabrous. Culms shallowly biconvex or rhombic in cross-section, 30–55 mm high, 0.8–1.4 mm wide, 1/2–2/3 length of longest leaves, pale yellow-brown at base, margins glabrous or scabrous with prickle hairs antrorse. Photosynthetic pathway ‘maximum cell distance count’ >1, C3.Inflorescence obovate in outline, a reduced panicle of 1–3 spikelets, 7–12 mm long, 2.5–5(–10) mm wide; involucral bract equal to or up to twice as long as the inflorescence, 9–24.5 mm long, sheath pink-red to dark red-brown at least proximally; rachis not flexous or reflexed. Spikelets 5–6.5 mm long, 1–2 mm diameter, with single bisexual flower; prophylls acute or emarginate, puberulous. Glumes 4, all of similar length; lowest 2 sterile, mucronate, outer face scabrous, with raised midrib; fertile glumes c. 6 mm long, apex subulate, midrib indistinct, outer face puberulous, margins glabrous. Perianth scales 6(–8), acute to acuminate, glabrous, 0.8–1.6 mm long. Anthers c. 1.4 mm long excluding apiculus; apiculus 0.4 mm long, glabrous. Nut elliptical in outline, pale green to mid brown depending on maturity, with 3 discolorous and raised ribs, c. 3 mm long, 1.4–1.7 mm diameter; style cap truncate, minutely colliculate.
Restricted to the South West, Central Highlands, West Coast and Mt Field regions of Tasmania; in alpine heath, herbfields, open forest and moorland at altitudes greater than about 700 m (Figure 6). At Tarn Shelf (Figure 4) and Mt Sprent this species grows in skeletal humus over or amongst rock outcrops, in epacrid–Proteaceae–Myrtaceae heath or Gymnoschoenus sphaerocephalus sedgeland.
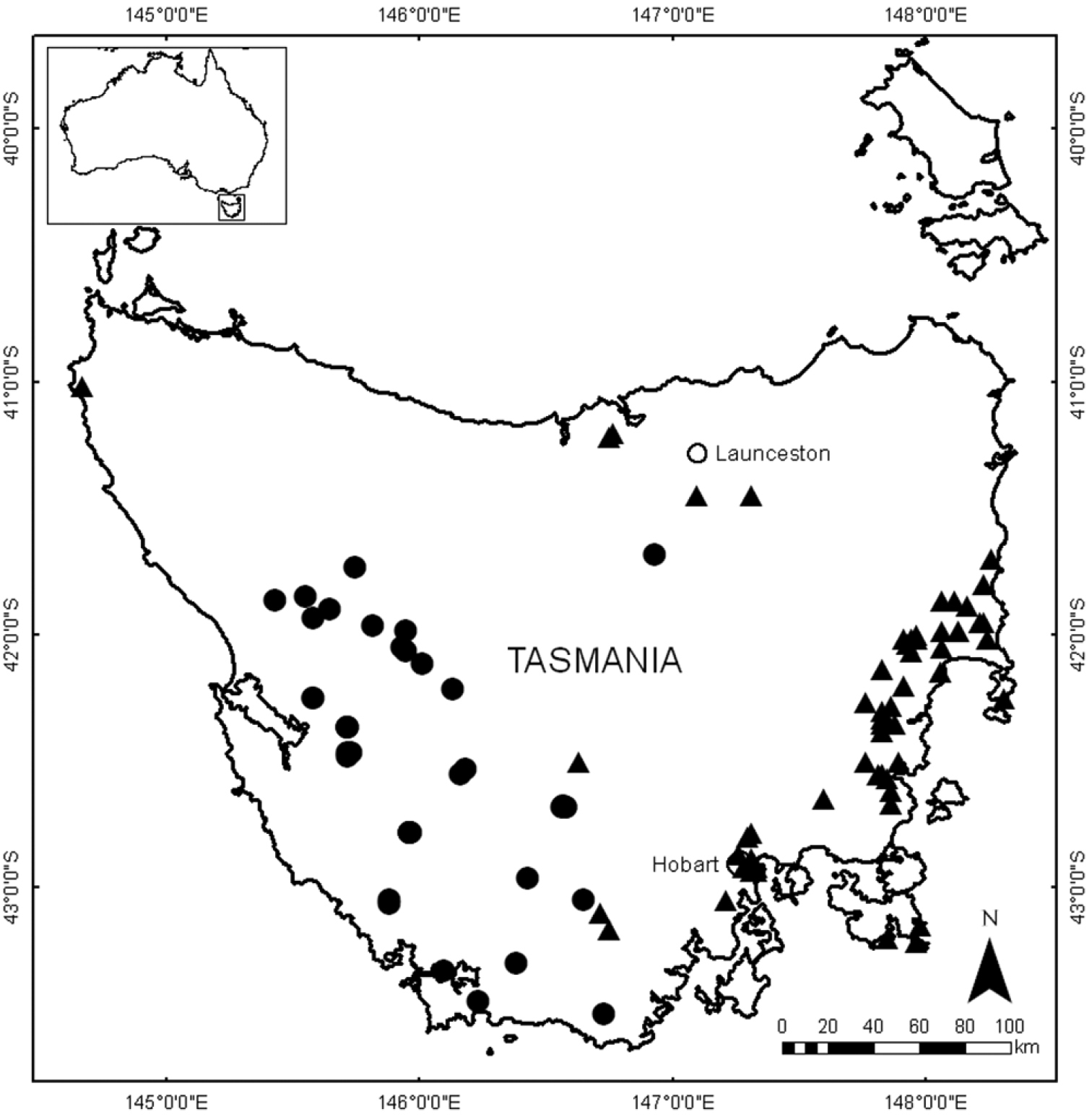
Distribution of Lepidosperma inops (solid black triangles) and Lepidosperma monticola (solid black circles) from herbarium specimen data. Cities indicated by open circles. All specimens seen by G.T. Plunkett (see Appendix 3 for vouchers).
On the basis of our current knowledge, this species would not warrant listing under either the International Union for Conservation of Nature Red List (
Named from the Latin mons, montis (mountain) and -cola (dweller), referring to the distribution of this species on the mountains of Central and South Western Tasmania.
AUSTRALIA. Tasmania: Central Highlands: 3 km SE of Pyramid Mountain, 1100 m, 14 Feb. 1983, A. Moscal 1776 (HO); Cradle plateau, 880 m, 7 Mar. 1949, W.M. Curtis s.n. (HO); High Dome, 24 Feb. 1994, J.B. Kirkpatrick s.n. (HO); Sticht Range, 20 km S of Tullah, 920 m, 16 Apr. 1990, P.A. Collier 4678 (HO). Mt Field: Shelf above University Hut, Lake Dobson, 1120 m, 1 Jan. 1949, W.M. Curtis s.n. (HO). South West: Elliot Range, summit, 900 m, 15 Jan. 1985, S.J. Jarman s.n. (HO, MEL, NSW); Mt Rugby, 1120 m, 16 Feb. 1978, S.J. Jarman s.n. (HO). West Coast: Mt Darwin, 1100 m, 8 Mar 1974, D.A. Ratkowsky s.n. (HO); Mt Dundas, summit, 860 m, 1 Mar. 1894, L. Rodway s.n. (HO); Range extending S to SE from Mt Curly, 5 Feb. 1985, S.J. Jarman 263 (HO).
Flowers November to February. Fruits December to April.
We thank HO and MEL for use of loan material, the staff at HO for use of facilities, access to specimens and specimen data and fieldwork tips, and NE for access to facilities and specimens. We acknowledge permission from National Parks and Wildlife in Tasmania to collect in areas under their management. Thanks also to Cate McGregor for mapping advice, Chrissie Prychid for assistance in the field and Ian Telford for constructive suggestions. GTP acknowledges receipt of an Australian Postgraduate Award (APA Scholarship), an Australian Biological Resources Study APATop-up grant, a UNE Botany Top-up Scholarship and funding from the School of Environmental and Rural Science.
Annotated character list used for phenetic analysis of species of Lepidosperma and description of Lepidosperma monticola sp. nov. Modified from DELTA ver. 1.04 database output.
Vegetative morphology:
#1. Rhizomes <whether long or short>/
1. long <shoots spreading>/
2. short <shoots clumped>/
• If long and short, both states scored
#2. Culms <length including inflorescence>/
mm long/
• Measured from base of plant (i.e. base of leaf sheaths) to the most distal inflorescence spikelet
#3. Culms <width at mid-third>/
mm wide/
• The greatest dimension in elliptical/flattened culms. Measured in the mid-third of culm length
#4. Culms <thickness at mid-third>/
mm thick/
#5. Culms width to thickness ratio <at mid-third>/
• Longest culm measured
#6. Culms <when dried; whether striate>/
1. striate/
2. not striate/
#7. Culms <whether deeply grooved>/
1. <deeply> grooved/
2. not <deeply> grooved/
• One or more deep grooves obvious at 10x magnification on fresh or dried material
#8. Culm margins <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
• Assessed along total length
#9. Culm margin hairs <orientation>/
1. antrorse/
2. retrorse/
3. divergent/
#10. Culm margin hairs <whether clumped>/
1. clumped/
2. evenly spaced/
#11. Culm margin hairs <whether embedded in resin>/
1. embedded in resin/
2. not embedded in resin/
#12. Culm margin hairs <position >/
1. restricted to the apex of culm/
2. along the upper third of culm/
3. continuous along culm/
4. along the lower third of culm/
5. along mid-third of culm/
#13. Culms <length relative to leaves>/
1. longer than leaves/
2. more or less equal to leaves/
3. shorter than leaves/
• Culm length includes inflorescence (see character 2)
#14. Culm bases <colour>/
1. green/
2. yellow-green/
3. pale yellow/
4. pale yellow-brown <straw-coloured>/
5. pale brown <pallid>/
6. orange-brown/
7. mid brown <tan>/
8. dark brown <dark chocolate>/
9. black-brown <intense dark brown>/
10. pink-brown/
11. red-brown/
12. red-pink/
13. dark red/
14. purplish/
15. black/
#15. Culm to leaf ratio <of lengths>/
• Relative lengths of longest culm and leaf
#16. Leaves <whether equitant>/
1. equitant/
2. not equitant/
#17. Leaves <angle of fan (ramet) in degrees;
degrees/
• Measured as the angle between the two outermost leaves on a single complete ramet/fan
#18. Leaves <longest leaf; length>/
mm long/
#19. Leaf sheaths <length>/
mm long/
#20. Leaf sheaths <colour>/
1. green/
2. yellow-green/
3. pale yellow/
4. pale yellow-brown <straw-coloured>/
5. pale brown <pallid>/
6. orange-brown/
7. mid brown <tan>/
8. dark brown <dark chocolate>/
9. black-brown <intense dark chocolate>/
10. pink-brown/
11. red-brown/
12. red-pink/
13. dark red/
14. purplish/
15. black/
• Assessed under dissecting microscope
#21. Leaf sheaths <whether resinous>/
1. resinous/
2. not resinous/
• Unopened sheath observed; comment on quantity
#22. Leaf sheath margins <opacity>/
1. translucent/
2. opaque/
#23. Leaf sheath margins <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
• Along margin towards apex
#24. Leaf sheath face <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#25. Ligule <apex shape>/
1. caudate/
2. accuminate/
3. acute/
4. obtuse/
5. rounded/
6. emarginate/
7. lobed/
8. truncate/
#26. Ligule <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
• Refers to the apex of the ligule.
#27. Leaf lamina <whether well-developed and photosynthetic at maturity>/
1. well-developed <photosynthetic, and at least some leaves shortly or distinctly laminate>/
2. reduced <functionally elaminate; leaves with lamina much reduced or absent>/
#28. Leaf lamina <whether rigid or flaccid>/
1. rigid/
2. flaccid/
#29. Leaf lamina <longest leaf; length>/
mm/
#30. Leaf lamina <width at mid-third of lamina length; longest leaf>/
mm wide/
#31. Leaf lamina <thickness in mid-third; not equivalent to width>/
mm thick/
#32. Leaf lamina <width to thickness ratio>/
• Measured in mid-third of lamina length
#33. Leaf lamina <general shape in cross-section>/
1. dorsiventrally flattened/
2. terete/
3. semiterete/
4. isobilaterally oblong/
5. isobilaterally flattened/
6. isobilaterally fusiform/
#34. Leaf lamina <colour of margins>/
1. with red margins/
2. with colourless margins/
3. with concolorous margins <i.e., margins same colour as faces>/
4. with brown margins/
5. with golden-orange margins/
#35. Leaf lamina <when dried; whether finely striate>/
1. striate/
2. not striate/
#36. Leaf lamina <whether deeply grooved>/
1. deeply grooved/
2. not deeply grooved/
#37. Leaf lamina margins <indumentum>/
1. glabrous/
2. scabrous/
3. papillose/
4. puberulous/
5. pilose/
• Assessed along whole length of lamina
#38. Leaf lamina margin hairs <orientation>/
1. antrorse/
2. retrorse/
3. divergent/
#39. Leaf lamina marginal hairs <whether clumped>/
1. clumped/
2. evenly spaced/
#40. Leaf lamina marginal hairs <whether embedded in resin>/
1. embedded in resin/
2. not embedded in resin/
#41. Leaf lamina marginal hairs <position >/
1. restricted to the lamina apex/
2. along the upper third of lamina/
3. continuous along lamina/
4. along the lower third of lamina/
5. along the lower half of lamina/
6. along mid-third of lamina/
7. along upper half of lamina/
#42. Leaf lamina to sheaths ratio <of lengths>/
Inflorescence morphology:
#43. Inflorescence <outline; i.e. overall plane shape>/
1. broadly ovate/
2. ovate/
3. narrowly ovate/
4. linear/
5. oblong/
6. narrowly oblong/
7. circular/
8. elliptic/
9. narrowly elliptic/
10. obovate/
11. narrowly obovate/
12. oblate/
13. fan-shaped/
14. narrowly rhombic/
15. broadly rhombic/
#44. Inflorescence <form>/
1. single spikelet/
2. panicle-like/
#45. Inflorescence <length>/
mm long/
• Measured from the base of the lowest primary inflorescence bract to tip of distal spikelet.
#46. Inflorescence <width at widest point>/
mm wide/
#47. Inflorescence <number of nodes on primary axis>/
noded/
• Score the maximum number per specimen; start with lowest node
#48. Inflorescence lowest internode <length along main axis>/
mm long/
• Measured from the base of lowest primary inflorescence bract to the base of the next sheath on the main inflorescence axis.
#49. Inflorescence lateral branches <number per node>/
per node/
• Scored for first lateral branch only
#50. Inflorescence lateral branches <whether elongated>/
1. elongated <branch visible above sheath of inflorescence bract>/
2. contracted <not visible beyond sheath>/
• Scored only where lateral branches with more than one spikelet were present.
#51. Inflorescence lateral branches <longest branch; length>/
mm long/
• Measured from lowest node to tip of the distal spikelet
#52. Proximal <inflorescence> lateral branch <length>/
mm long/
#53. Proximal <inflorescence> lateral branch <width>/
mm wide/
#54. Involucral bract <total length>/
mm/
#55. Involucral bract to inflorescence ratio <of total lengths>/
#56. Involucral bract sheath <length>/
mm/
#57. Involucral bract sheath <colour>/
1. green/
2. yellow-green/
3. pale yellow/
4. pale yellow-brown
5. pale brown/
6. orange-brown/
7. mid brown/
8. dark brown/
9. black-brown/
10. pink-brown/
11. red-brown/
12. red-pink/
13. dark red/
14. purplish/
15. black/
16. grey/
#58. Involucral bract sheath <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#59. Involucral bract sheath apex <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#60. Involucral bract sheath margin <opacity>/
1. translucent/
2. opaque/
#61. Involucral bract sheath ligule <whether with a free limb>/
1. with a free limb/
2. without a free limb/
#62. Involucral bract sheath ligule <apex shape>/
1. caudate/
2. acuminate/
3. acute/
4. obtuse/
5. emarginate/
6. lobed/
7. rounded/
8. truncate/
#63. Involucral bract sheath ligule <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#64. Involucral bract lamina to sheath ratio <of lengths>/
#65. Involucral bract <whether with a well-developed lamina>/
1. lamina well-developed <photosynthetic>/
2. reduced <non-photosynthetic at maturity>/
• Lamina well-developd or much narrower than sheath or mucro-like or absent
#66. Involucral bract lamina <length>/
mm long/
#67. Involucral bract lamina <width>/
mm wide/
#68. Involucral bract lamina <shape in cross-section>/
1. isobilaterally flattened <or elliptical> along entire length <of lamina>/
2. triangular at lamina apex <but becoming isobilaterally elliptical above ligule; e.g. Lepidosperma tortuosum>/
3. triangular along entire length <of lamina>/
4. triangular at lamina apex <and becoming half-rhombic towards base>/
5. triangular at lamina apex <and becoming trullate towards base>/
6. triangular at lamina apex <and becoming circular towards base>/
7. circular along entire length/
8. dorsiventrally flattened along entire length/
9. dorsiventrally flattened at apex <and becoming oblong at base>/
10. dorsiventrally crescentiform at apex <and becoming isobilaterally elliptical towards base>/
#69. Involucral bract lamina <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#70. Involucral bract lamina apex <shape>/
1. aristate/
2. caudate/
3. subulate/
4. acuminate/
5. acute/
6. obtuse/
7. rounded/
#71. Rachis <whether flexuous>/
1. flexuous/
2. not flexous/
• Spikelet-bearing axis of inflorescence.
#72. Rachis <whether reflexed>/
1. reflexed <e.g. Lepidosperma tortuosum>/
2. not reflexed/
Spikelet and floral morphology:
#73. Spikelet prophyll <shape of apex>/
1. obtuse/
2. truncate/
3. emarginate/
4. acute/
#74. Spikelet prophyll margin <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#75. Spikelet prophylls outside <abaxial> face <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#76. Spikelet prophyll keel <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#77. Spikelets <number per inflorescence>/
1. less than 30/
2. more than 30/
#78. Spikelets <number per lowest lateral branch>/
per lowest lateral branch/
#79. Spikelets <length excluding any pedicel>/
mm long/
• Measure spikelet from base to apex
#80. Spikelets <diameter>/
mm diameter/
#81. Sterile floral bract apex <shape>/
1. aristate/
2. subulate/
3. acuminate/
4. acute/
5. obtuse/
6. rounded/
7. truncate/
8. emarginate/
#82. Sterile floral bract midrib <whether raised>/
1. raised/
2. flush/
#83. Sterile floral bract <whether mucronate>/
1. mucronate/
2. muticous <not mucronate>/
#84. Sterile floral bract outside (abaxial) face <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#85. Sterile floral bract margins <form of margin indumentum>/
1. glabrous/
2. scabrous/
3. puberulose/
4. pilose/
#86. Sterile floral bracts <number per spikelet>/
per spikelet/
#87. Fertile floral bracts <number per spikelet>/
per spikelet/
#88. Fertile female floral bract apex <shape>/
1. aristate/
2. subulate/
3. acuminate/
4. acute/
5. obtuse/
6. rounded/
#89. Fertile floral bracts <whether mucronate>/
1. mucronate/
2. muticous <not mucronate>/
#90. Fertile floral bract midrib <whether obvious at 10x>/
1. apparent/
2. indistinct/
#91. Fertile floral bract midrib <indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#92. Fertile floral bracts outside face <indumentum; abaxially, excluding midrib and margins>/
1. glabrous/
2. scabrous/
3. papillose/
4. puberulous/
5. pilose/
#93. Fertile floral bracts inside face <indumentum; adaxially, excluding margins>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#94. Fertile floral bracts margins <form of margin indumentum>/
1. glabrous/
2. scabrous/
3. puberulous/
4. pilose/
#95. Fertile floral bract nerves <number visible with 10x magnification>/
#96. Female-fertile flowers <includes bisexual flowers; number>/
per spikelet/
#97. Male flowers <number>/
per spikelet/
#98. Female-fertile floral bracts <length>/
mm long/
#99. Female-fertile floral bracts <width; measured in si tu>/
mm wide/
#100. Total number of flowers per spikelet/
#101. Stamens <number per flower>/
per flower/
#102. Anther <length excluding apiculus>/
mm long excl. apiculus/
#103. Anthers <width>/
mm wide/
#104. Anther apiculus <length>/
mm long/
#105. Anther apiculus <indumentum>/
1. glabrous/
2. scabrous/
#106. Style branches <number>/
per pistil/
Fruit and perianth morphology:
#107. Perianth scale <number>/
#108. Perianth scale <shape>/
1. narrow ovate/
2. ovate/
3. broadly ovate/
4. circular/
#109. Perianth scale apex <shape>/
1. caudate <drawn into long tail>/
2. subulate/
3. acuminate/
4. acute/
5. rounded/
6. obtuse/
#110. Perianth scale apex <indumentum>/
1. glabrous/
2. indumented/
#111. Perianth scale apex hairs <position>/
1. restricted to the very tip/
2. not restricted to the tip <and extending proximally>/
#112. Perianth scale fused base <maximum length>/
mm/
#113. Perianth scale free limb <maximum length>/
mm/
#114. Perianth scale <width>/
mm/
• Measured at widest part of scale
#115. Perianth scales <whether overlapping at the base >/
1. overlapping/
2. not overlapping/
#116. Fruit <length including perianth>/
mm long including perianth/
#117. Fruit <length excluding perianth>/
mm long excluding perianth/
#118. Fruit <diameter>/
mm diameter/
• At widest diameter
#119. Fruit <shape in broadest lateral view>/
1. spherical/
2. ovate <widest near base>/
3. oblong <with parallel sides>/
4. elliptical <widest in middle >/
5. obovate <widest near apex>
#120. Fruit ribs <whether distinct from fruit faces>/
1. discolorous with faces/
2. concolorous with faces/
#121. Fruit ribs <whether raised>/
1. raised <above face of fruit>/
2. not raised <ribs more or less flush with surface of nut>/
3. sunken <below face of fruit>/
#122. Fruit ribs <colour>/
1. green/
2. pale green/
3. pale yellow/
4. yellow-brown/
5. pale brown/
6. orange-brown/
7. mid brown/
8. dark brown/
9. red-brown/
10. black/
#123. Fruit face <colour at maturity>/
1. green/
2. pale green/
3. pale brown-green/
4. yellow-green/
5. pale yellow/
6. yellow-brown/
7. pale brown/
8. orange-brown/
9. mid brown/
10. dark brown/
11. red-brown/
12. black/
#124. Fruit faces <whether wrinkled at maturity>/
1. wrinkled/
2. not wrinkled/
#125. Fruit style cap <length>/
mm/
#126. Fruit style cap apex <shape>/
1. truncate/
2. rounded/
3. obtuse/
4. acute/
#127. Fruit style cap <colour>/
1. green/
2. pale green/
3. pale brown-green/
4. yellow-green/
5. pale yellow/
6. yellow-brown/
7. pale brown/
8. orange-brown/
9. mid brown/
10. dark brown/
11. red-brown/
12. black/
• Record if fruit immature, recently mature or well mature.
#128. Fruit style cap <indumentum>/
1. glabrous/
2. scabrous/
3. papillous/
4. <minutely> colliculate/
#129. Fruit style base <presence fused on top of style cap>/
1. present/
2. absent/
#130. Fruit style base <length>/
mm/
#131. Fruit style base <width>/
mm/
Culm anatomy
• As seen in cross-section. Many characters are from or modified from
#132. Culms <shape in cross-section>/
1. triangular/
2. circular/
3. truncated circular/
4. subhemispherical/
5. square/
6. trullate or rhombic/
7. crescentiform/
8. fusiform/
9. broadly elliptical/
10. elliptical/
11. narrowly elliptical/
12. ovate/
13. ‘almond-shaped’<ovate with one margin acute and the other obtuse or rounded>/
14. oblong/
15. shallowly hexagonal/
16. ‘Swiss-escutcheon-shaped’<shaped like a Swiss escutcheon, which is an elaborated heart shape with two convex sides and a truncate apex with a short projection in the middle >/
#133. Culm margins <shape in cross-section>/
1. rounded <e.g. in Lepidosperma viscidum>/
2. obtuse/
3. acute/
4. acuminate/
#134. Culm stomata <position relative to epidermal cells>/
1. raised/
2. flush/
3. sunken/
#135. Culm vascular bundles <number>/
#136. Culm vascular bundles <number of distinct size classes>/
#137. Culm vascular bundles enclosed within chlorenchyma <presence>/
1. present/
2. absent/
• Individual vascular bundles surrounded by chlorenchyma cells
#138. Culm vascular bundles enclosed within parenchyma <presence>/
1. present/
2. absent/
• Individual vascular bundles surrounded by parenchyma cells
#139. Culm vascular bundles between chlorenchyma and parenchyma <presence>/
1. present/
2. absent/
• Individual vascular bundles may be positioned between, and contacting both, chlorenchyma and parenchyma, i.e. not completely surrounded by one tissue type.
#140. Culm sclerenchymatous fibre caps <presence>/
1. present/
2. absent/
#141. Culm sclerenchymatous fibre girders <presence>/
1. present/
2. absent/
• Girders connecting vascular bundles and epidermis.
#142. Culm sclerenchymatous fibre girders between vascular bundles <presence>/
1. present <e.g. in Lepidosperma concavum (J.J. Bruhl 25 74)>/
2. absent/
#143. Culm sclerenchymatous fibre strands <presence>/
1. present/
2. absent/
#144. Culm sclerenchymatous fibre strands <whether all contacting epidermis>/
1. all contacting epidermis/
2. some not contacting epidermis/
• Some small strands occur which are surrounded by mesophyll or parenchyma and do not contact the epidermis or vascular bundles.
#145. Culm pith or parenchyma <whether homogeneous or heterogeneous>/
1. homogeneous/
2. heterogeneous/
• Homogeneous culm pith /parenchyma consists mainly of a single, uniform tissue type. Heterogeneous pith is composed of two distinct tissue types: (1) tissue where the cells have relatively thick cell-walls, usually associated with the vascular bundles; and (2) ‘translucent tissue’ where the cells are generally larger and have thin cell-walls.
#146. Culm pith or parenchyma <heterogeneous morphology>/
1. vascular bundle ‘rings’ surrounding translucent tissue/
2. large amount of pith with thin ‘bridges’ between opposite vascular bundles/
#147. Culm multicellular trichomes <whether present>/
1. absent/
2. present singularly/
3. present in groups/
#148. Culm epidermal zones <relative width>/
1. wider costally/
2. equal in width/
3. wider intercostally/
#149. Culm epidermal cell silica bodies <presence>/
1. present <within thin-walled epidermal cells>/
2. absent <and epidermal cells thick walled>/
#150. Culm epidermal cell silica bodies <distribution>/
1. throughout epidermis/
2. restricted to margin epidermis/
#151. Culm epidermal ‘silica cells’ <position relative to non-silica epidermal cells>/
1. raised/
2. flush/
3. sunken/
• ‘Silica cells’ are epidermal cells containing one or more silica bodies
#152. Culm sub-epidermal sclerenchymatous strands and/or girders width <mean number of cells>/
• Mean of counts of five adjacent strands and/or girders. Within each strand and/or girder count the number of sclerenchymatous cells adjacent to the epidermis as seen in cross-section. Counted in mid-third of section if possible.
#153. Culm sub-epidermal sclerenchymatous strands and/or girders length <mean number of cells>/
• Mean count of five adjacent strands and/or girders. Within each strand and/or girder count the number of sclerenchymatous cells from its inner end to the epidermis, not including the mestome sheath. Counted in mid-third of section where possible.
Dataset of Lepidosperma species from DELTA file. Operational taxonomic unit (OTU) code in first column. Characters marked with * not included in phenetic analysis. See Table 1 for OTU details and Appendix 1 for character list.
| Character | curBag01 | curCon01 | curMac01 | curRos01 | eldCou01 | eldEld01 | eldFie01 | eldFie02 | eldMcC01 | eldPro01 | eldSpr01 | eldSpr02 | eldSpr03 | inoBre01 | inoGra01 | inoLen01 | inoUni01 | inoUni02 | torBaro02 | torBarT01 | torBla01 | torBoo01 | torDem01 | torMul01 | torWer01 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1* | 2 | 1/2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| 2 | 99 | 116 | 93 | 185 | 31 | 43 | 51 | 35 | 48 | 41 | 51 | 55 | 39 | 49 | 17.5 | 45 | 45 | 56 | 341 | 233 | 423 | 350 | 319 | 343 | 502 |
| 3 | 1.23 | 1.34 | 1.21 | 1.21 | 1.18 | 1.23 | 0.8 | 0.81 | 0.97 | 1.3 | 1.4 | 1.24 | 1.31 | 0.74 | 0.83 | 1.02 | 0.72 | 0.67 | 1.44 | 0.83 | 0.82 | 1.55 | 1.21 | 1.07 | 0.82 |
| 4 | 0.39 | 0.41 | 0.72 | 0.65 | 0.36 | 0.44 | 0.36 | 0.39 | 0.46 | 0.27 | 0.51 | 0.28 | 0.39 | 0.42 | 0.47 | 0.41 | 0.35 | 0.38 | 0.62 | 0.61 | 0.6 | 0.77 | 0.55 | 0.45 | 0.66 |
| 5 | 3.15 | 3.27 | 1.68 | 1.86 | 3.28 | 2.8 | 2.22 | 2.08 | 2.11 | 4.81 | 2.75 | 4.43 | 3.36 | 1.76 | 1.77 | 2.49 | 2.06 | 1.76 | 2.32 | 1.36 | 1.37 | 2.01 | 2.2 | 2.38 | 1.24 |
| 6* | 2 | 1 | 2 | 1/2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1/2 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 7* | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1/2 | 2 | 1/2 | 1/2 | 1/2 | 2 | 2 |
| 8 | 2 | 2 | 2 | 2 | 2 | 1&2 | 1&2 | 1/2 | 2 | 2 | 1/2 | 1/2 | 1/2 | 1 | 1 | 1/2 | 1/2 | 1/2 | 2 | 2 | 2 | 2 | 2 | 2 | 1/2 |
| 9 | 3 | 3 | 3 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 3 | 3 | 1/3 | 1 | 3 | 3 | 3 | ||
| 10 | 2 | 2 | 2 | 2 | 2 | 1 | 1/2 | 1/2 | 1/2 | 2 | 1/2 | 2 | 2 | 1 | 2 | 2 | 1/2 | 1 | 1 | 2 | 2 | 1/2 | |||
| 11* | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | ||
| 12 | 3 | 3 | 3 | 2& 3/5 | 3 | 3 | 2/4 | 1/3 | 3 | 3 | 3 | 2/3 | 2/3 | 2 | 1 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | ||
| 13 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | 2 | 1 | 1 | 1 | 1 | 1 |
| 14* | 3/5 | 3 | 3/5 | 3/5 | 2/3 | 3/4 | 3/5 | 3 | 2/4 | 1/3-4 | 3 | 3 | 3 | 3 | 3 | 3/5-7 | 3 | 3 | 3 | 7-11-13 | 11/12 | 3 | 2/3/5 | 3/5 | 3/12 |
| 15 | 0.36 | 0.47 | 0.54 | 0.82 | 0.56 | 0.59 | 0.58 | 0.7 | 0.66 | 0.53 | 0.57 | 0.69 | 0.6 | 0.35 | 0.27 | 0.34 | 0.37 | 0.31 | 1.45 | 1.16 | 3.38 | 1.66 | 1.29 | 1.66 | 1.56 |
| 16* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 17 | 10 | 10 | 15 | 6 | 22 | 20 | 20 | 20 | 23 | 24 | 26 | 27 | 25 | 12 | 15 | 15 | 15 | 9 | 7 | 5 | 5 | 8 | 5 | 6 | 7 |
| 18 | 272 | 246 | 173 | 225 | 55 | 73 | 88 | 50 | 73 | 77 | 79 | 80 | 65 | 139 | 65 | 133 | 121 | 181 | 235 | 227 | 125 | 211 | 247 | 207 | 321 |
| 19 | 40 | 34 | 25 | 26 | 16 | 22 | 25 | 12 | 14 | 23.5 | 22 | 22 | 12 | 29 | 15 | 30 | 19.5 | 36.5 | 35 | 26 | 50 | 25 | 23 | 20 | 31 |
| 20 | 3/5 | 4/7 | 3/5 | 4/5 | 3/4-5/12 | 3/12 | 1/3-4/5-12 | 3/5-12 | 3/4-5/7-12 | 5/7-12 | 3/5-7/12 | 3/5-7/12 | 2/3-4/5-7/11 | 3/5 | 4/5-7 | 4/5-7 | 3/5-7 | 1/5-7 | 3/4-5/7 | 2/4-5/7-12 | 8/11 /12 |
3/4-5 | 3/4 | 3/4-5 | 3/7-11/12 |
| 21 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 1/2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| 22* | 1/2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 23* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 24* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||||
| 25 | 4/6 | 4/5 | 4/6 | 3/4 | 2/3 | 1/2 | 2 | 3 | 2 | 1/2 | 2/3 | 2 | 2 | 2 | 2/3 | 2/3 | 1 | 1 | 6 | 6 | 6 | 4/6 | 8 | 6 | 6 |
| 26* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 27* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 28* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 29 | 232 | 212 | 148 | 199 | 39 | 51 | 63 | 38 | 59 | 63.5 | 68 | 58 | 53 | 110 | 50 | 103 | 101.5 | 144.5 | 200 | 201 | 75 | 86 | 224 | 187 | 279 |
| 30 | 1.4 | 1.43 | 1.76 | 1.18 | 1.15 | 1.24 | 1.15 | 0.99 | 1.01 | 1.31 | 1.55 | 1.44 | 1.46 | 0.71 | 1.06 | 0.95 | 0.88 | 1.2 | 1 | 0.63 | 0.63 | 1.32 | 1.39 | 1.22 | 0.77 |
| 31 | 0.4 | 0.4 | 0.52 | 0.49 | 0.32 | 0.32 | 0.31 | 0.35 | 0.21 | 0.29 | 0.48 | 0.29 | 0.3 | 0.19 | 0.22 | 0.34 | 0.18 | 0.22 | 0.33 | 0.36 | 0.39 | 0.44 | 0.34 | 0.31 | 0.5 |
| 32 | 3.5 | 3.57 | 3.38 | 2.41 | 3.59 | 3.87 | 3.71 | 2.83 | 4.81 | 4.52 | 3.23 | 4.97 | 4.87 | 3.74 | 4.82 | 2.79 | 4.89 | 5.45 | 3.03 | 1.75 | 1.62 | 3 | 4.09 | 3.94 | 1.54 |
| 33* | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| 34 | 2/3 | 2/3 | 3 | 3 | 2/3 | 3 | 2/3 | 3 | 3 | 2/3 | 2/3 | 2/3 | 2/3 | 3 | 3 | 3 | 3 | 3 | 2/3 | 2/3 | 3 | 2 | 2/3 | 2/3 | 3 |
| 35* | 2 | 1 | 2 | 1/2 | 1 | 1 | 1 | 1 | 1 | 1 | 1/2 | 1 | 1 | 1 | 1 | 1/2 | 1 | 1 | 1 | 1/2 | 1 | 1 | 1 | 1 | 1 |
| 36 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 1/2 | 1 | 1 | 1 | 1/2 |
| 37 | 2 | 2 | 2 | 2 | 1/2 | 2 | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 | 1/2-4 | 1/2 | 2 | 1/2 | 1/2 | 2 | 2 | 2 | 2 | 1/2 |
| 38 | 1/3 | 3 | 1/3 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 3 | 3 | 1 | 1/3 | 3 | 1/3 | 1 |
| 39 | 2 | 2 | 1 | 2 | 1 | 1/2 | 1 | 1 | 1/2 | 1/2 | 1/2 | 2 | 2 | 1/2 | 1/2 | 1/2 | 1 | 2 | 1/2 | 1 | 1 | 2 | 1/2 | 1 | |
| 40* | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |
| 41 | 3 | 3 | 3 | 3 | 1/2 | 2/3 | 2/3 | 1/2 | 1/2 | 1 | 1/2/6 | 1/6 | 2/3 | 7 | 7 | 7 | 1 | 3 | 3 | 1 | 3 | 3 | 3 | 3 | |
| 42 | 5.8 | 6.24 | 5.92 | 7.65 | 2.44 | 2.32 | 2.52 | 3.17 | 4.21 | 2.28 | 3.09 | 2.64 | 4.42 | 3.79 | 3.33 | 3.43 | 5.21 | 3.96 | 5.71 | 7.73 | 1.5 | 3.44 | 9.74 | 9.35 | 6.64 |
| 43 | 9 | 2/3/ 8/9 |
6/9 | 2/8 | 10 | 10 | 2 | 2 | 10 | 10 | 1/2 | 2/10 | 1/2 | 3 | 3 | 3 | 3 | 3 | 2/5 | 1/8 | 2/5/8 | 1/5 | 2/8 | 2 | 1/12 |
| 44 | 2 | 2 | 2 | 2 | 2 | 2 | 1/2 | 1 | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 | 2 | 1/2 | 1/2 | 1/2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| 45 | 30 | 29 | 30 | 35 | 7 | 9.5 | 12 | 9 | 10 | 10.5 | 8.5 | 8.5 | 9 | 10 | 5.5 | 12 | 11 | 7.7 | 12 | 9 | 12 | 11 | 7.5 | 11 | 11 |
| 46 | 10 | 7 | 8 | 13 | 3.5 | 2.5 | 5 | 3 | 10 | 5 | 2.5 | 4 | 5.5 | 3.5 | 4 | 7 | 3.5 | 4.4 | 12 | 12 | 9 | 8 | 10 | 11.5 | 10.5 |
| 47 | 7 | 8 | 7 | 7 | 2 | 3 | 2 | 1 | 3 | 3 | 2 | 2 | 2 | 2 | 2 | 2/3 | 2 | 3 | 3 | 3 | 4 | 4 | 4 | 3 | 5 |
| 48 | 8 | 8 | 10.5 | 9.5 | 1 | 3.5 | 2.5 | 2 | 2.5 | 2 | 2.5 | 4.5 | 2 | 6.5 | 5.5 | 2 | 4 | 2 | 7 | 4 | 2.5 | 3 | 3 | ||
| 49* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| 50 | 1/2 | 1/2 | 1/2 | 1/2 | 2 | 1 | 2 | 2 | 2 | ||||||||||||||||
| 51 | 16 | 16 | 15 | 22 | 6.5 | 5 | 7 | 8 | 8.5 | 7.5 | 5 | 6.5 | 6 | 5 | 7.5 | 6 | 6.1 | 12.5 | 9 | 12 | 11 | 9.5 | 11 | 11 | |
| 52 | 16 | 16 | 15 | 22 | 6.5 | 5 | 7 | 8 | 8.5 | 7.5 | 5 | 6.5 | 6 | 5 | 7.5 | 6 | 6.1 | 12.5 | 9 | 12 | 11 | 9.5 | 11 | 11 | |
| 53 | 4 | 2.5 | 3 | 5 | 1.5 | 1 | 2 | 2 | 1.5 | 1 | 1.5 | 2 | 1.5 | 2 | 4 | 2 | 1.5 | 7.5 | 3 | 7 | 8 | 5 | 2.5 | 7 | |
| 54 | 19 | 2.5 | 21.5 | 22 | 9 | 12 | 20.5 | 13 | 10.5 | 24.5 | 12 | 9.5 | 10.5 | 52 | 27 | 59 | 49 | 41 | 13 | 9.5 | 28 | 11 | 8 | 9.5 | 10 |
| 55 | 0.63 | 31 | 0.72 | 0.63 | 1.29 | 1.26 | 1.71 | 1.44 | 1.05 | 2.33 | 1.41 | 1.12 | 1.17 | 5.2 | 4.91 | 4.92 | 4.45 | 5.32 | 1.08 | 1.06 | 2.33 | 1 | 1.07 | 0.86 | 0.91 |
| 56 | 10 | 1.07 | 10.5 | 10 | 5 | 5.5 | 7.5 | 7 | 5.5 | 8 | 7 | 5.5 | 5.5 | 10 | 6.5 | 9 | 7.5 | 5.5 | 3 | 2 | 4 | 4 | 2 | 2.5 | 2.5 |
| 57 | 5/7-16 | 5 | 5/7 | 5/7-8 | 7/11 /13 |
5/7-11 | 5/7-8 | 5/7-8/9-11 | 7/8-11/13 | 5/7-8/12 | 5/7 | 7/8 | 7/8 | 2 | 1/3-4 | 3/5-7 | 3/4-5 | 1/3-5/7 | 4/7-11 | 7/11 | 11/13 | 3/7-8/12 | 7/8 | 7/8 | 8/11 |
| 58 | 2 | 2 | 2 | 2/3 | 2 | 2 | 1 | 1 | 2 | 1/2 | 1 | 1 | 1 | 2 | 1 | 2/3 | 2 | 2 | 1 | 2 | 1 | 3 | 2 | 2 | 1 |
| 59 | 2 | 2 | 1/2 | 1/2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 60 | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1/2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 |
| 61* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| 62 | 3/4 | 4 | 5 | 2/3 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 1 | 1 | 5 | 5 | 5 | 5 | 5 | 5/8 | 5 | |
| 63 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| 64 | 0.9 | 2.44 | 1.05 | 1.2 | 0.8 | 1.18 | 0.52 | 0.57 | 0.91 | 2.06 | 1.4 | 0.73 | 0.91 | 4.2 | 3.15 | 5.56 | 5.53 | 6.45 | 3.33 | 3.75 | 6 | 2.25 | 3 | 0.74 | 3 |
| 65* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 66 | 9 | 22 | 11 | 10 | 4 | 6.5 | 13 | 6 | 5 | 16.5 | 5 | 4 | 5 | 42 | 20.5 | 50 | 41.5 | 35.5 | 10 | 7.2 | 24 | 9 | 6 | 7 | 7.5 |
| 67 | 0.88 | 1.04 | 0.75 | 1.1 | 0.72 | 0.65 | 0.89 | 0.56 | 0.61 | 1.24 | 0.43 | 0.74 | 0.74 | 0.85 | 1 | 1.11 | 0.65 | 0.82 | 0.97 | 0.72 | 0.73 | 0.95 | 0.68 | 0.51 | 0.51 |
| 68 | 2/3 | 2/3 | 2/3 | 2/3 | 2/10 | 10 | 10 | 2 | 10 | 2/10 | 2/10 | 2 | 2/10 | 2 | 2 | 2 | 2 | 2 | 2 | 1/2/3 | 2 | 2 | 2 | 2/3 | 2 |
| 69 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1/2 | 2/3 | 2 | 2 | 1/2 | 2 | 2 | 2/3 | 2 | 2 | 1/2 |
| 70 | 4 | 2/4 | 4 | 5 | 5/7 | 2 | 7 | 5/6/7 | 4 | 7 | 7 | 7 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 3/4 | 3/4 | 4 | 4 | 5/3 | |
| 71* | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 2 | 1/2 | 1/2 | ||||
| 72 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| 73 | 3 | 4 | 3 | 3 | 4 | 3 | 4 | 3/4 | 3/4 | 3/4 | 2/4 | 4 | 4 | 4 | 3 | 1 | 3 | 3 | 3/4 | 3/4 | |||||
| 74 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | 3 | 1 | 3 | 2 | 1 | 1 | 3 | 1 | 1 | 3 | 3 | 1 | 1 | 1 | ||||
| 75 | 1 | 1 | 1 | 1 | 1/2 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||
| 76 | 3 | 3 | 3 | 3 | 3/4 | 3 | 3 | 3/4 | 3 | 3 | 3 | 3 | 3 | 3 | 2 | 3 | 3/4 | 2/3 | 3 | 3 | |||||
| 77* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||
| 78 | 4 | 5 | 4 | 6 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1/2 | 1 | 1 | 1 | 3 | 1 | 3 | 3 | 2 | 1 | 4 | ||
| 79 | 6.5 | 6 | 3.6 | 6.3 | 6.5 | 5 | 6.5 | 6.5 | 6.5 | 6.5 | 5 | 5.4 | 4.8 | 4.5 | 5.3 | 5.6 | 7.8 | 6 | 5.3 | 6.8 | 6.8 | 6.1 | 6.7 | ||
| 80 | 1.6 | 1.2 | 1.6 | 1.7 | 1.5 | 1 | 2 | 2 | 1.6 | 1.7 | 1.5 | 1.3 | 2.3 | 2 | 2 | 1.5 | 2.6 | 1.5 | 1.2 | 1.9 | 1.6 | 1.35 | 1.4 | ||
| 81* | 1 | 1 | 1 | 1 | 2 | 1/2 | 1 | 1 | 8 | 7/8 | 7 | 1 | 8 | 8 | |||||||||||
| 82* | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | |||||||||||
| 83* | 1 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||
| 84* | 2 | 3 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 2 | 2 | 1 | 1/2 | 1/2 | |||||||||||
| 85* | 3 | 3 | 3 | 1 | 2 | 3 | 1 | 2 | 1 | 3 | 3 | 1 | 1 | 1/2 | |||||||||||
| 86* | 2 | 3 | 3 | 2 | 3 | 3 | 3 | 3 | 1 | 3 | 2 | 3 | 5 | ||||||||||||
| 87* | 1 | 3 | 2 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 2 | 2 | 3 | ||||||||||||
| 88* | 3 | 2 | 2 | 2 | 2/3 | 2 | 2 | 3 | 2 | 4 | 3 | 2 | 3 | 2 | |||||||||||
| 89* | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 1 | 2 | 2 | 2 | |||||||||||
| 90* | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 2 | |||||||||||
| 91* | 2 | 2 | 3 | 3 | 2 | 2 | 3 | 1 | 2 | 3 | 3 | 1/2 | 3 | 2 | |||||||||||
| 92* | 2 | 2 | 4 | 4 | 2 | 2 | 4 | 2 | 2 | 4 | 4 | 1/2 | 4 | 2 | |||||||||||
| 93* | 1 | 2 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||
| 94* | 3 | 3 | 3 | 1 | 3 | 3 | 3 | 2 | 1/2 | 3 | 3 | 1 | 3 | 2 | |||||||||||
| 95* | 1 | 1 | 2 | 0 | 1 | 2 | 3 | 0 | 1 | 1 | 1 | 1 | 2 | 0 | |||||||||||
| 96* | 1 | 3 | 2 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | ||||||||||||
| 97* | 1 | 3 | 2 | 1 | 1 | 1 | 2 | 2 | 2 | 1 | 2 | 2 | 3 | ||||||||||||
| 98* | 3.7 | 3.4 | 5.4 | 5.7 | 3.7 | 4.8 | 3.1 | 2.5 | 6 | 4.4 | 5 | 4.4 | 5.05 | ||||||||||||
| 99* | 0.9 | 0.6 | 0.5 | 1 | 0.8 | 1.25 | 0.9 | 4 | 1.1 | 0.9 | 0.9 | 0.6 | 0.8 | ||||||||||||
| 100* | 1 | 3 | 2 | 1 | 1 | 2 | 2 | 2 | 2 | 1 | 2 | 2 | 3 | ||||||||||||
| 101* | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |||||||||||||||
| 102* | 0.9 | 1.6 | 1.4 | 1.5 | 1.15 | 1.7 | 1.8 | ||||||||||||||||||
| 103* | 0.35 | 0.3 | 3.5 | 0.2 | 0.25 | 0.3 | 0.2 | ||||||||||||||||||
| 104* | 0.3 | 0.5 | 0.4 | 0.45 | 0.2 | 0.2 | 0.2 | ||||||||||||||||||
| 105* | 2 | 1 | 1 | 2 | 1 | 1/2 | |||||||||||||||||||
| 106* | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | |||||||||||||||||
| 107 | 6/7 | 6 | 6 | 7 | 6 | 8 | 6 | 6 | 8 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | ||||||||
| 108* | 1 | 1 | 1 | 1 | 1 | 1 | 1/2 | 1 | 1 | 1 | 1 | 1/2 | 1 | 1/2 | 2 | 1 | 1/2 | ||||||||
| 109* | 3/4 | 2 | 2/3 | 3 | 3/4 | 2/3 | 3 | 3 | 3 | 3 | 2/3 | 2/3 | 3/4 | 3 | 3/4-5 | 3 | 2/3 | ||||||||
| 110* | 1/2 | 1 | 1/2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |||||||||||
| 111* | 1 | 1 | |||||||||||||||||||||||
| 112 | 0.4 | 0.2 | 0.05 | 0.3 | 0.1 | 0.4 | 0.3 | 0.3 | 0.1 | 0.3 | 0.1 | 0.2 | 0.2 | 0.2 | 0.1 | 0.35 | 0.4 | ||||||||
| 113 | 1.1 | 0.8 | 1.05 | 1.3 | 1.1 | 1.6 | 0.8 | 1.5 | 1.1 | 1.5 | 0.9 | 0.8 | 0.9 | 0.8 | 0.9 | 0.6 | 1.2 | ||||||||
| 114 | 0.4 | 0.5 | 0.5 | 0.4 | 0.4 | 0.5 | 0.6 | 0.5 | 0.6 | 0.4 | 0.8 | 0.5 | 0.35 | 0.5 | 5.5 | 0.3 | 0.5 | ||||||||
| 115* | 1 | 2 | 1 | 1/2 | 1/2 | 1 | 1/2 | 2 | 1 | 2 | 1 | 2 | 2 | 1 | 1 | 2 | |||||||||
| 116 | 3.6 | 3 | 2.65 | 3.1 | 3 | 3.4 | 3.1 | 2.95 | 3.05 | 3.1 | 3.15 | 2.7 | 2.8 | 2.5 | 2.8 | 2.6 | 2.8 | ||||||||
| 117 | 3.2 | 2.8 | 2.6 | 2.8 | 2.9 | 3 | 2.8 | 2.65 | 2.95 | 2.8 | 3.05 | 2.5 | 2.6 | 2.3 | 2.7 | 2 | 2.6 | ||||||||
| 118 | 1.5 | 1.6 | 1.35 | 1.7 | 1.65 | 1.7 | 1.6 | 1.4 | 1.5 | 1.45 | 1.7 | 1.4 | 1.45 | 1.5 | 1.5 | 1.4 | 1.45 | ||||||||
| 119* | 4 | 4 | 4 | 5 | 4 | 4 | 4 | 4 | 4 | 4/5 | 4 | 4 | 2/4 | 4 | 4 | 4 | 4 | ||||||||
| 120 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||
| 121 | 1 | 1/2 | 1 | 1 | 1 | 2 | 2 | 1 | 2 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||
| 122* | 3/5 | 3/5 | 5 | 5 | 1/3 | 5 | 3/5 | 2 | 6 | 2 | 7/8 | 3 | 3 | 3/5 | 3 | 3/5 | 3/4 | ||||||||
| 123* | 1/3 | 2/7-9 | 9 | 7/9 | 3/9 | 9 | 9/10 | 4 | 3 | 2/7 | 10 | 7 | 1/7 | 3 | 3 | 1 | 3/9 | ||||||||
| 124* | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 1 | 2 | 2 | 2 | 1/2 | ||||||||
| 125 | 0.3 | 0.2 | 0.3 | 0.4 | 0.3 | 0.25 | 0.6 | 0.2 | 0.2 | 0.3 | 3 | 0.4 | 0.4 | 0.3 | 0.3 | 0.3 | 0.4 | ||||||||
| 126* | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1/2 | 3 | 1/3 | 1 | ||||||||||
| 127* | 10 | 5/9 | 7 | 7 | 5/10 | 8 | 10/12 | 2/5 | 10/12 | 7/9 | 12 | 11 | 7 | 5 | 6 | ||||||||||
| 128 | 1 | 1 | 1 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||
| 129* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | |||||||||
| 130 | 0.1 | 0.05 | 0.05 | 0.1 | 0.2 | 0.2 | 0.13 | 0.2 | 0.1 | 1.5 | 0.05 | 0.1 | 0.1 | 0.1 | 0.1 | ||||||||||
| 131 | 0.1 | 0.1 | 0.15 | 0.15 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 2 | 0.1 | 0.15 | 0.1 | 0.1 | 0.15 | ||||||||||
| 132 | 10/11 | 10 | 11 | 11 | 13 | 6 | 13 | 13 | 13 | 13 | 13 | 10 | 10/13 | 14 | 9/14 | 10 | 9/12 | 2/9 | 13 | 16 | 15 | 4 | 15 | 15 | 16 |
| 133 | 3/4 | 4 | 3/4 | 2/3 | 1&3 | 1&3 | 1/3 | 1/4 | 1&4 | 1&4 | 2/3 | 2/3 | 2/3 | 1 | 1 | 3/1 | 2&4 | 1&2 | 3&4 | 3 | 4 | 1/2 | |||
| 134* | 2 | 1/2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |
| 135 | 32 | 38 | 29 | 23 | 13 | 13 | 10 | 10 | 10 | 10 | 11 | 12 | 15 | 13 | 11 | 9 | 10 | 12 | 26 | 17 | 18 | 31 | 21 | 2 | 19 |
| 136 | 3 | 2 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | 3 |
| 137* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 138 | 1 | 1 | 1 | 2 | 2 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 2 | 2 | 2 | 2 | 2 | 1 | 2 | 2 | 1 | 2 | 2 | |
| 139 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| 140* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 |
| 141 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| 142* | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |
| 143* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 144* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| 145 | 2 | 2 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| 146 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||||
| 147 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 |
| 148 | 1 | 1 | 2/3 | 1/2 | 3 | 2 | 1/2 | 2 | 3 | 3 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1/2 | 1 | 2 | 2 | 1 | 2 |
| 149 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| 150* | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||
| 151 | 2 | 2 | 2 | 2 | 2/3 | 2/3 | 2 | 2 | 3 | 2/3 | 2 | 2/3 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | ||||
| 152 | 8 | 7 | 7.2 | 7.8 | 7.8 | 5.8 | 6.6 | 9 | 5.6 | 6 | 7 | 7.2 | 5.4 | 8.2 | 6.2 | 6.8 | 17 | 8 | 7.8 | 5.8 | 8.2 | 7 | 7.4 | 5.8 | 8 |
| 153 | 12.8 | 14 | 10.8 | 12.6 | 6.2 | 4.8 | 3.4 | 3.2 | 5 | 4.4 | 4.6 | 4.6 | 5 | 8 | 12.2 | 5.4 | 3.8 | 6 | 7.2 | 5.6 | 9.8 | 9.2 | 8.8 | 7.6 | 9.6 |
Index to numbered collections of Lepidosperma seen by G.T. Plunkett
All specimens are in HO unless otherwise indicated.
Allan, M. s.n. 30 Sept. 1975 HO 135425, s.n. 19 Oct. 1979 HO 515594 (Lepidosperma inops).
Beauglehole, A.C. 25084 MEL (Lepidosperma curtisiae).
Brown, M.J. 1365 (Lepidosperma monticola).
Bruhl, J.J. 630 (Lepidosperma monticola); 2407 NE (Lepidosperma inops); 2453 NE (Lepidosperma tortuosum); 3005 NE (Lepidosperma curtisiae).
Buchanan, A.M. 3293, 3018a, 7276, 7629 (Lepidosperma inops); 9981 (Lepidosperma monticola); 10880 (Lepidosperma inops); 12360 (Lepidosperma monticola); 13324, 14710, 15288 (Lepidosperma inops).
Coates, F. s.n. 25 Jan. 1990 HO 127069 (Lepidosperma inops).
Collier, P.A. 945, 1015, 1391, 1488, 1500, 1628, 1900, 2338, 2522, 2560, 3247 (Lepidosperma inops); 3844 (Lepidosperma monticola); 4016, 4105, 4354 (Lepidosperma inops); 4405, 4678 (Lepidosperma monticola), 5254 (Lepidosperma inops).
Curtis, W.M. s.n. 1 Jan. 1949 HO 135483 (Lepidosperma inops); s.n. 1 Jan. 1949 HO 135412, s.n. 1 Jan. 1949 HO 135486, s.n. 1 Jan. 1949 HO 135489, s.n. 7 Mar. 1949 HO 135487 (Lepidosperma monticola); s.n. 10 Sept. 1951 HO 135488, s.n. 3 Dec. 1972 HO 24548, s.n. 28 Oct. 1978 HO 135485, s.n. 28 Oct. 1978 HO 534984 (Lepidosperma inops).
Davies, F.E. 1241 (Lepidosperma inops).
Dobson, A.T. 77290 (Lepidosperma monticola).
Duncan, F. 2, 1120, s.n. 22 Mar. 1994 HO 317251, s.n. 1 Aug. 1995 HO 314343, s.n. 1 Aug. 1995 HO 314344, s.n. 31 Aug. 1995 HO 314345 (Lepidosperma inops).
Griffith, S.J. s.n. 20 Apr. 1993 NE (Lepidosperma tortuosum).
Hodgon, J. 391c NE, 415 NE (Lepidosperma tortuosum).
Hosking, J.R. 3041 NE (Lepidosperma tortuosum).
Hunter, J.T. 5068 NE (Lepidosperma tortuosum).
Jackson, W.D. s.n. 1 Apr. 1960 HO 24549, 30 Aug. 1983 HO 68636 (Lepidosperma inops).
Jarman, S.J. 157, 180, 235, 263, s.n. 16 Feb. 1978 HO 411020, s.n. 20 Dec 1978 HO31334, s.n. 13 Dec. 1979 HO 411752 (Lepidosperma monticola); s.n. 12 Aug. 1980 HO 319561 (Lepidosperma inops); s.n. 8 Feb. 1984 HO 412112, s.n. 21 Mar. 1984 HO 412037, s.n. 15 Jan.1985 HO 443183, s.n. 4 Mar. 1986 HO 412044, s.n. 12 Mar. 1986 HO 411078, s.n. 10 Apr. 1986 HO 412047, s.n. date unknown HO 411443 (Lepidosperma monticola).
Klaphake, V. s.n. 1 Jan. 1998 NE (Lepidosperma tortuosum)
Moscal, A. 214, 397, 409 (Lepidosperma inops); 1776, 1795, 4636 (Lepidosperma monticola); 6182 NE (Lepidosperma inops); 8022, 8039 (Lepidosperma monticola); 8040 (Lepidosperma curtisiae); 8254, 8285, 8291, 8363 (Lepidosperma monticola); 9441, 18031 NE (Lepidosperma inops); 19548, 28626 (Lepidosperma monticola).
North, A.J. s.n. 7 Oct. 1992 HO 132741, s.n. 7 Oct. 1992 HO 132754 (Lepidosperma inops); s.n. 8 Oct. 1996 HO 322622 NE (Lepidosperma curtisiae); s.n. 26 Sept. 2004 HO 530824 (Lepidosperma inops).
Plunkett, G.T. 94a, 94b, 95, 96, 98, 99, 100 (Lepidosperma monticola); 112, 115 (Lepidosperma inops) all NE.
Rao, C.V. s.n. 1 Jan. 1956 HO 135544 (Lepidosperma inops).
Ratkowsky, D.A. 545, 1082 NE, s.n. 1 Jan. 1974 HO 135490 (Lepidosperma inops); 8 Mar. 1974 HO 135415 (Lepidosperma monticola).
Rodway, L. s.n. 1 Mar. 1892 HO 26449 (Lepidosperma inops); s.n. 1 Mar. 1894 HO 26450 (Lepidosperma monticola).
Smith, J.M.B. 342 (Lepidosperma monticola).
Williams, K.J. s.n. 18 May 1987 HO 531508 (Lepidosperma inops).
Wilson, K.L. 6522, 6662 (Lepidosperma inops).
Ziegeler, D. 84, 114 (Lepidosperma inops).