





(C) 2012 Neil Snow. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Five new species of Rhodamnia are proposed for New Guinea, including Rhodamnia asekiensis, Rhodamnia daymanensis, Rhodamnia makumak, Rhodamnia taratot, and Rhodamnia waigeoensis. Rhodamnia sharpeana, known previously only in Australia, is reported for the first time for Papua New Guinea. Detailed species descriptions and associated taxonomic data are provided for all species. A key is provided for species of Rhodamnia with stellate trichomes. Given the overall paucity of collections, all species are tentatively assigned as Data Deficient following IUCN conservation recommendations.
Australia, conservation, Myrtaceae, New Guinea, new species, Rhodamnia, systematics
Rhodamnia Jack is recognized easily among the baccate genera of Myrtaceae in Malesia and Melanesia by its 4-merous flowers (apart from a 5-merous species in New Caledonia), uniloclular ovaries, parietal placentation, and sclerotic seed coats (
Based on the author’s current taxonomic perpectives and including those newly proposed here, Rhodamnia includes 42 species. The genus occurs from Myanmar, Thailand and China through Malesia and Australia, and east through Melanesia to the Solomon Islands (
The five species newly proposed here were recognized during curatorial duties associated with floristic inventories in Papua New Guinea by Bishop Museum. The purpose of this paper is to describe the new species of Rhodamnia, summarize their diagnostic character traits, and to provide a distribution map and conservation threat assessments. It also discusses the first New Guinean occurrences of Rhodamnia sharpeana, previously known only from Australia (
Measurements are based primarily on dried herbarium specimens, although dimensions for flowers and fruits were supplemented by rehydrating material in boiling water. Terminology follows that used in recent treatments for the genus (
The data underpinning the new species described in this paper are deposited at GBIF, the Global Biodiversity Information Facility, http://ipt.pensoft.net/ipt/resource.do?r=snow_1_rhodamnia_new_guinea.
urn:lsid:ipni.org:names:77123882-1
http://species-id.net/wiki/Rhodamnia_asekiensis
Figures 1, 2Resembling Rhodamnia latifolia but with distinctly larger leaves that have an acute to acuminate apex and differing by the larger fruits.
Papua New Guinea. Morobe Province, Aseki, Menyama Subdistrict, 7°21'S, 146°10'E, 20 May 1968, H. Streimann & A. Kairo NGF 39049(holotype: BISH [sheet no. 30914]!; isotypes: A!, CANB!, K!, L!, LAE n.v., NSW n.v., US!)
Trees of unknown height; crown dense. Bark of main bole light grey, vertically fissured. Indumentum (branchlets, flowers, fruit) short-sericeous, sparsely to moderately dense, color more or less saffron (
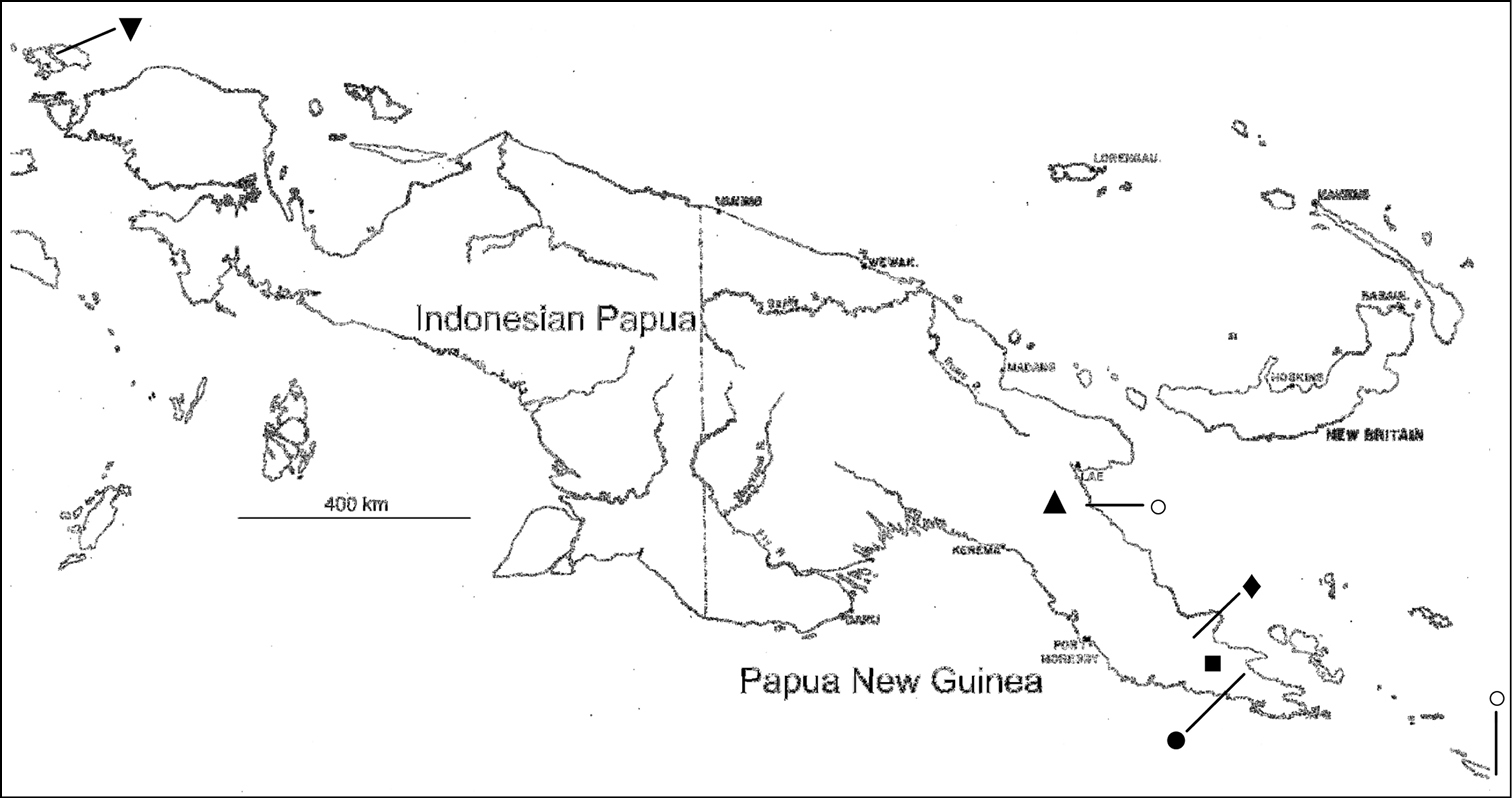
Island of New Guinea, showing localities of new species. Triangle (▲) = Rhodamnia asekiensis; Square (■) = Rhodamnia daymanensis; Diamond (♦) = Rhodamnia makumak (at bottom of line); Closed circle (●) = Rhodamnia toratot (at top of line); Inverted triangle (▼) = Rhodamnia waigeoensis (upper left, at bottom of line); Open circles (○) = Rhodamnia sharpeana (left end of line [upper] and bottom of line [lower, on truncated eastern half of Tagula Island]).

Rhodamnia asekiensis N. Snow. Photo of the holotype at BISH (H. Streimann and A. Kairo NGF 39049).
Flowering unknown; fruiting confirmed only for May.
Known only from Morobe Province, Papua New Guinea, on the side of a ridge in a secondary forest; ca. 1465 m.
Data Deficient; but the subsequent lack of collections of this species over the past 45 years locally and regionally suggest that Threatened might more accurately reflect is true status.
Rhodamnia asekiensis is included among the “pearly” group of species (
urn:lsid:ipni.org:names:77123883-1
http://species-id.net/wiki/Rhodamnia_daymanensis
Figures 1, 3Resembling Rhodamnia lancifolia but differing by its more deeply sulcate petiole, broader leaves, and shorter yellowish indumentum on the abaxial laminar surface.
PapUa New Guinea. Milne Bay District, north slopes of Mt. Dayman, Maneau Range, 2250 m, ca. 9°47'S, 149°18'E, 2 Jun 1953, L. J. Brass 22718 (holotype: A!; isotype: L!)
Trees 15–18 m. Indumentum (branchlets, inflorescence axis, flowers) densely appressed sericeous or sericeous-villous (trichomes yellowish). Branchlets terete to compressed, brown (dried), epidermis smooth, becoming flakey or scaly, sericeous-villous. Leaves opposite, evenly distributed along branchlets, strongly discolorous, internodes 1–3 cm long; venation perfect basal acrodromous, secondary and tertiary veins visible above and below; intramarginal vein closely paralleling leaf margin, 0.5–0.7 mm from margin at midpoint of blade. Colleters absent. Petioles 5–9 mm long, slightly sulcate throughout. Leaf blades 3.8–7.0 cm long, (1.1–)1.5–3.0(–3.5) cm wide, elliptic (rarely broadly elliptic), base cuneate, apex narrowly acuminate, tip acute and somewhat falcate; adaxial surface matte, sericeous but becoming glabrescent, midvein slightly sulcate in proximal half but more or less flush distally; abaxial surface densely sericeous between the secondary and tertiary veins, midvein projecting throughout, oil glands (if present) entirely obscured by indumentum. Inflorescence axillary, flowers solitary (=monads) or in 3-flowered cymes (=botryoids), solitary to paired or fascicled in axils, pedicels of monads up to 1–5 mm long, rigid and ascending. Bracteoles 1.5–3.0 mm long, less than 0.5 mm wide at base, linear, mostly erect or ascending, mostly persistent in flower. Hypanthium 2.5–3.3 mm long, ca. 2.5 mm wide at base of calyx lobes, cupulate, densely hairy. Calyx lobes 2.7–3.0 mm long, broadly obtuse, glabrescent adaxially, densely sericeous abaxially. Petals 5.5–7.0 mm long, 3.0–3.5 mm wide, ovate to narrowly ovate, whitish, mostly glabrous adaxially, densely sericeous abaxially, oil glands common. Stamens 65–75, multiseriate; staminal disk short-hairy; filaments 2–3 mm long; anther sacs 0.3–0.5 mm long, globose to subcylindrical, sub-basifixed or basifixed, crowned by a single large apical gland. Style ca. 4.5 mm long, glabrous; stigma narrow to slightly capitate, prominently papillose. Ovary and locule 1, placentas 2, linear, ovules disposed in regular rows.

Rhodamnia daymanensis N. Snow. Photo of the holotype at A (L. J. Brass 22718).
Flowering in June; fruiting interval unknown.
Papua New Guinea, Milne Bay Province, north slopes of Mt. Dayman in the Maneau Range; mossy forest of ridge crests over metamorphic rocks (see
Data Deficient. The collection label indicates the species was common (at least locally) at the time of its collection in 1953. However, the absence of additional collections over the past sixty years suggests that Threatened might more accurately reflect its true status.
Rhodamnia daymanensis appears to be part of the “pearly” group (
Among the species of Rhodamnia in New Guinea occurring at elevations above 2000 meters with a similar abaxial indumentum, Rhodamnia daymanensis mostly closely resembles Rhodamnia lancifolia. The type collection of Rhodamnia lancifolia (2425 m) is approximately 25 km west of the type locality of Rhodamnia daymanensis (2250 m) in similar habitats. However, Rhodamnia lancifolia differs by its more narrowly elliptic leaves, a less pronounced petiolar sulcus, slightly impressed adaxial laminar midvein (vs. more deeply impressed in Rhodamnia daymanensis in the proximal half), and the more yellowish (and longer, on average) abaxial laminar indumentum. The adaxial leaf surface of Rhodamnia lancifolia, for which there are many collections, is nearly black when dried, which contrasts with the fucosus (dark greyish brown,
urn:lsid:ipni.org:names:77123884-1
http://species-id.net/wiki/Rhodamnia_makumak
Figures 1, 4Resembling other species of Rhodamnia having stellate trichomes but differing by its sessile to subsessile flowers and narrowly elliptic leaves with occasionally falcate apices.
Papua New Guinea. Milne Bay Province: E. of Mt. Suckling in valley of the upper Maiyu R[iver] c. 15 km WNW of Biniguni airstrip, ca. 9°40'S, 149°10'E, ca. 350 m, 7 Jul 1972, R. Pullen 8433(holotype: A! [no accession numer]; isotypes: BO n.v., BRI!, CANB!, L!, LAE n.v., K!, TNS n.v.).
Trees to 25 m. Buttresses present but low of stature; fluting or twisting absent. Bark of main trunk reticulate-flaky, brownish. Indumentum, where present (branchlets, petioles, abaxial leaf surface, distal portion of adaxial leaf midvein, peduncles, bracteoles, hypanthium, calyx lobes, adaxial petal surfaces), densely tomentose and velvety in texture, consisting of stellate, ferrugineous trichomes. Branchlets terete to compressed. Leaves opposite, more or less evenly distributed along branchlets, discolorous; venation perfect or imperfect suprabasal acrodromous, secondary and tertiary veins faint but visible adaxially, intramarginal vein faintly visible from adaxially, tracing irregularly between tips of secondary veins and ca 0.5 mm from blade margin. Colleters absent. Petioles 4.5–6.5 mm long, rounded in transverse section. Leaf blades 4.5–7.5 cm long, 1.4–2.2 cm wide, narrowly elliptic, base cuneate, margin flat, apex acuminate and sometimes falcate, tip (uppermost 10% of blade) acute; adaxial surface matte, midvein slightly sulcate more or less throughout to sometimes flush distally, tomentose proximally; abaxial surface orangish-velvety by virtue of indumentum, midvein projecting throughout. Inflorescence terminal and lateral, solitary or paired to mostly a fasciculate cluster of monads, the monads sessile or on pedicels up to 3 mm long. Bracteoles 1.8–2.3 mm long, 0.4–0.6 mm wide, linear, rigid, ascending to erect, the apex not reaching base of calyx lobes, persisting. Hypanthium campanulate; anthopodium (if present) up to 1 mm long; metaxyphylls absent. Calyx lobes 4, 2.2–2.7 mm long, 2 (of the 4 lobes) more or less rectangular (length-width ratio 3:2), slightly longer than the 2 shorter, broadly ovate (3:2) lobes; adaxial surface densely tomentose or somewhat less so basally and near margins, abaxial surface densely tomentose throughout. Petals (material sparse) 2–2.5 mm long, 2–2.3 mm wide, elliptic to ovate, tomentose above and below. Stamens ca. 30–40, filaments 2–3 mm long; anther sacs ca. 0.5 mm long, globose, sub-basifixed. Style 3.5–4 mm long, hairy below; stigma narrow to slightly capitate. Fruit not seen.
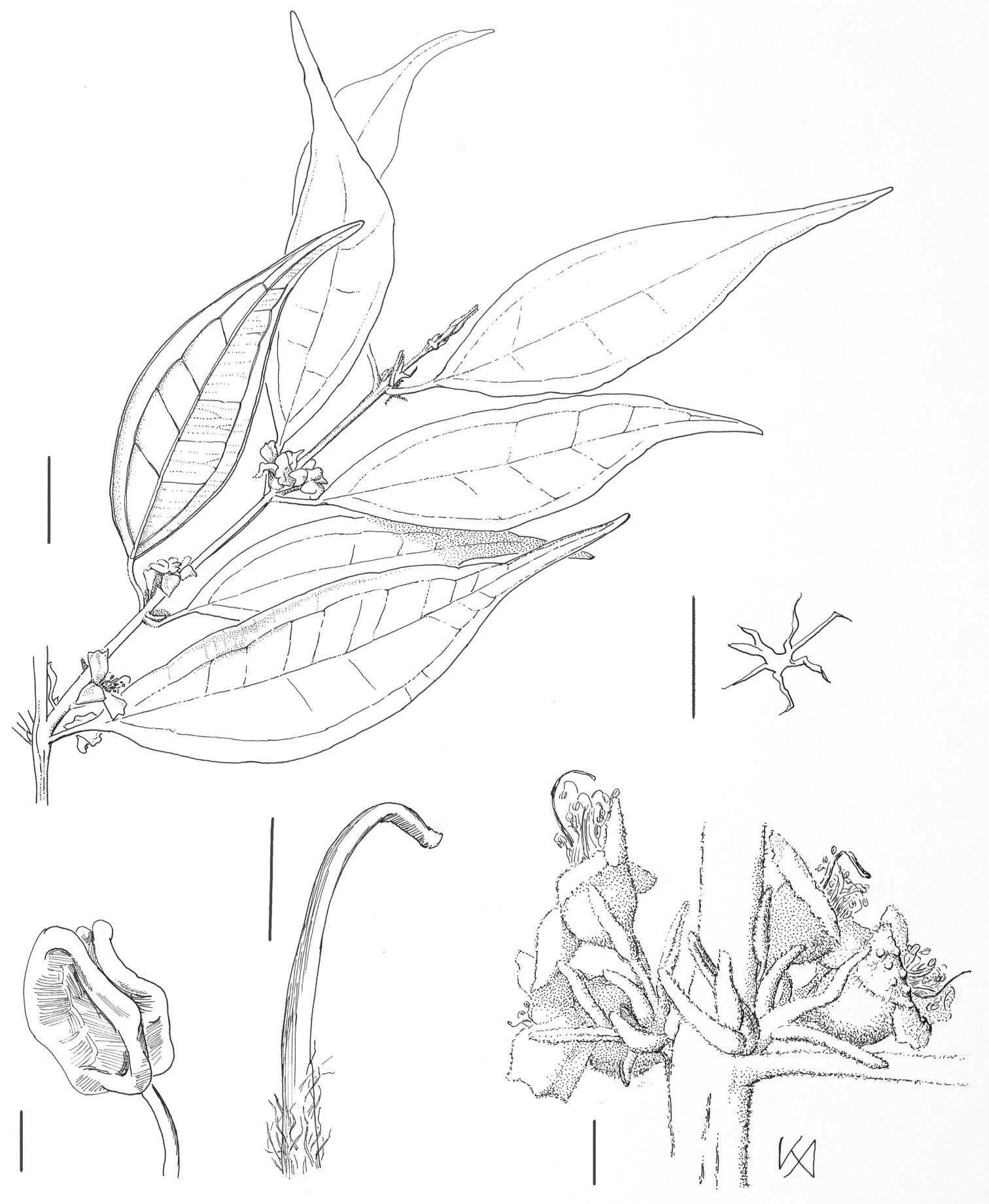
Rhodamnia makumak N. Snow. Drawing of isotype at A (R. Pullen 8433). Clockwise from top: branchlet (bar = 1 cm); stellate trichome (bar = 0.1 mm); inflorescences (bar = 1 mm); style (bar = 1 mm); dehisced anther (bar = 0.3 mm).
Flowering confirmed only for early July but likely also in late June; fruiting unknown but probably June to July and possibly longer.
Papua New Guinea, Milne Bay Province; known only from rainforest on a plateau of ca. 350 meters elevation.
Data Deficient.
From the local vernacular name “makumak” as a noun in the nominative.
Makumak in the local Daga language.
Rhodamnia makumak ispart of a group of species characterized by a hypothesized synapomorphy of stellate trichomes (
Rhodamnia makumak is said to be a large tree with low buttresses (dimensions of buttresses lacking on label). The best diagnostic characters include the stellate, ferrugineous indumentum on leaves and hypanthium, which imparts a densely velvety appearance; the sessile to subsessile axillary clusters of flowers; and narrowly elliptic leaves bearing an acuminate and sometimes falcate apex. A provisional key to species with stellate trichomes follows.
Key to species of Rhodamnia with stellate trichomes
| 1 | Leaves elliptic to broadly elliptic, 11–18 cm long, apex abruptly cuspidate-caudate | Rhodamnia kamialiensis N. Snow & W. N. Takeuchi |
| – | Leaves narrowly elliptic or narrowly ovate to ovate or elliptic, 4–12 cm long, apex acute to acuminate | 2 |
| 2 | Flowers pedicellate, pedicels mostly > 5 mm long; leaf apex acute | Rhodamnia propinqua C.T. White |
| – | Flowers sessile or nearly so; leaf apex acute to acuminate | 3 |
| 3 | Flowers sessile or pedicels to 3 mm; leaf apex acuminate and sometimes falcate | Rhodamnia makumak |
| – | Flowers pedicillate, pedicels 3.5–6 mm long; leaf apex acute to acuminate, rarely mucronate, never falcate | 4 |
| 4 | Seeds with a thin but pronounced equatorial ridge; plants of Australia, 650–1300 m | Rhodamnia blairiana F. Muell |
| – | Seeds lacking an equatorial ridge; plants of Australia and Papua New Guinea, sea level to ca. 500 m | Rhodamnia sharpeana N. Snow |
urn:lsid:ipni.org:names:77123885-1
http://species-id.net/wiki/Rhodamnia_toratot
Figures 1, 5Superficially resembling species with stellate and typically ferrugineous trichomes but differing by its highly crispate trichomes.
Papua New Guinea. Milne Bay District, Nowata, c. 6 miles W. of Rabaraba, 09°59'S, 149°43'E, ca. 520 m, 5 Jul 1969, R. Pullen 7709 (holotype: K!; isotypes: A! [barcode 00307479], BISH!, BO n.v., BRI!, CANB n.v. [00217141.1 and 00217141.2], G!, L n.v., LAE n.v.).
Trees to ca. 5.5 m. Bark of main bole unknown. Indumentum (branchlets, leaves, flowers, fruit) mostly densely tomentose-lanate, the trichomes highly crisped, ferrugineous, and generally somewhat appressed (see also description of abaxial leaf surface below). Branchlets terete to slightly compressed, reddish-brown (dried); epidermis smooth but finely and evenly striate throughout becoming somewhat fissured with age; oil glands sparse to common (obscured by indumentum on younger branchlets). Leaves opposite, evenly distributed along branchlets, somewhat discolorous; venation perfect basal to slightly suprabasal perfect or imperfect acrodromous, secondary and higher order veins abaxially prominent, the secondaries varying greatly in prominence (and thus hard to estimate numerically) but mostly spaced (2–)3–7 mm along the midvein; intramarginal vein less pronounced than the secondary veins, paralleling leaf margin closely, mostly ca. 0.5–1.0 mm from margin at midpoint of blade. Colleters absent. Petioles 9.0–13.5 mm long, terete, densely lanate-tomentose. Leaf blades 6.5–12.0 cm long, 3.1–5.2 cm wide, elliptic to ovate, base cuneate to nearly rounded, apex acute to acuminate, tip acute to acuminate; adaxial surface matte, initially lanate but becoming glabrescent, midvein slightly impressed throughout; abaxial surface lanate along midvein and secondary veins when younger, increasingly glabrous with age, densely/minutely hoary between veins, midvein raised prominently throughout. Inflorescence lateral in current season’s growth, flowers solitary to mostly densely fascicled, sessile to subsessile with pedicels up to 5 mm long, the pedicels lax, sometimes bending. Bracteoles ca. 2.0–2.5 mm long, ca. 0.5 mm wide at base, linear, persisting in flower and frequently in fruit. Hypanthium cupulate. Calyx lobes 2.5–3.5 mm long, ovate, apex obtuse, densely hairy abaxially but adaxially less so (especially proximally) with age, persisent and erect in fruit. Petals (material scanty), ca. 4.0–4.5 mm long, up to 3.5 mm wide, broadly obovate, more or less glabrous adaxially, densely lanate abaxially. Staminal disk ca. 4.5 mm in diameter; staminal ring narrow, shortly villous-lanate (trichomes whitish-yellow). Stamens numerous (estimated 75–105); anthers sacs (material scanty) cylindrical, ca. 1.0 mm long, bearing a single apical gland; filament length unknown. Styles not seen, but persisting bases densely lanate in fruit. Ovary with 1 locule; placentas 2; placentation parietal; ovules numerous. Fruit subglobose, 7.5–8.5 mm long (probably immature) x 8.0–9.5 mm wide, greenish when young but becoming brownish on account of dense indumentum. Seeds somewhat compressed, ca. 1–2 mm thick.

Rhodamnia toratot N. Snow. Drawing of isotype at BISH (R. Pullen 7709). Clockwise from top center: branchlet; detail of branchlet indumentum; bracteole; abaxial leaf surface; seed. Scale bars = 1 mm, except branchlet bar = 1 cm.
Flowering unknown; fruiting in July.
Milne Bay Province in Papua New Guinea; in secondary (regrowth) forest with Dodonaea Adans. (Sapindaceae) and Castanopsis (D. Don) Spach (Fagaceae).
Data Deficient; possibly Threatened for same reasons cited above for Rhodamnia asekiensis.
The specific epithet is derived from toratot as a noun in the nominative.
Locally known as toratot in the Nowata language.
The tomentose to lanate indumentum on the branchlets and inflorescences (Fig. 5) of Rhodamnia toratot suggests its inclusion in the “villous” group of species (
urn:lsid:ipni.org:names:77123886-1
http://species-id.net/wiki/Rhodamnia_waigeoensis
Figures 1, 6Closely resembling but differing from Rhodamnia novoguineensis by its thicker and more rigid pedicels, thickly coriaceous leaves, basal acrodromous venation, densely yellowish abaxial laminar indumentum, and solitary flowers.
Indonesia. Waigeo Island, Go Isthmus, path from Poean Bay to Fofak Bay, 17 Feb 1955, P. van Royen 5556(holotype: A! [bar code no. 00307477]; isotypes: CANB!, K!, L n.v.).
Trees 5–7 m; girth to 15 cm. Branchlets terete to slightly compressed, the epidermis later becoming fissured; indumentum densely sericeous, mostly yellowish or somewhat ferrugineous but becoming more whitish with age. Leaves opposite, evenly distributed along branchlets, discolorous, glossy above and below, the nacreous sheen below imparted by the dense, tightly appressed greenish-white indumentum. Colleters absent. Petioles 5–6.5 mm long, somewhat flattened above, densely sericeous (or somewhat tomentose with age), the indumentum yellowish but aging whitish. Leaf blades (3.5–)6.0–10.0 cm long, (1.8–)2.5–3.7 cm wide, narrowly ovate to ovate, surface flat or slightly wavy; base cuneate, apex acuminate and occasionally somewhat falcate, tip acute; venation perfect basal acrodromous; secondary veins numerous but thin, ca. 0.8–2.0 mm apart; marginal nerve prominent, mostly 0.7–0.9 mm from mid-leaf margins; margins flat; adaxial surface sparsely sericeous, midvein flush throughout, oil glands invisible; abaxial surface densely sericeous with greenish-whitish indumentum but this mostly not obscuring venation, midvein raised throughout, oil glands invisible. Inflorescence (limited material) a 3-flowered cyme, terminal, solitary (one per leaf subtending leaf); peduncle ca. 5 mm, stiff, terete in transsection, densely yellowish-orangish sericeous; pedicel to ca. 3 mm long, indumentum as per peduncle. Bracteoles 2, narrowly triangular and stiffly erect, ca. 2 mm long, ca. 0.5 mm wide, sericeous, sometimes persisting into fruit. Hypanthium cupulate, densely sericeous, oil glands absent, texture smooth. Calyx lobes 4, 2.3–3.5 mm long, broadly ovate, sericeous above, densely sericeous below, more or less reflexed in fruit. Petals 4, 5.5–7.0 mm long, width uncertain (material scanty), apparently obovate to broadly obovate, white (based on specimen label), sparsely sericeous above, densely sericeous below. Staminal disk 2.5–3.5 mm wide, densely short-hairy. Ovary apex densely short-hairy. Stamen number uncertain but almost certainly greater than 20, filaments and anthers red (from specimen label); anthers subcylindrical (material scanty), ca. 0.5 mm. Stigma not seen. Locule 1, placentation parietal, placentas 2, ovules numerous. Fruit (reportedly immature) globose-subglobse, up to 8 mm long and 9.5 mm wide, light green when immature, densely sericeous but indumentum thinning with maturity. Seeds irregularly angular, up to 4 mm long (small sample), up to 9 per fruit, crowded; seed coat hard. Embryos not seen.

Rhodamnia waigeoensis N. Snow. Photo of the the holotype at A (P. van Royen 5556)
Flowering February; fruiting in January and February.
Waigeo Island, Indonesia; from ca. 10–150 m elevation in xerophytic, Myrtaceae-dominated vegetation at lower elevations behind and upslope of the village of Waifoi, and from transitional forests dominated by Decaspermum J.R. Forst. & G. Forst. (Myrtaceae) or Vatica rassak Blume(=Vatica papuana Schum. & Hollr. [synonym]) (Dipterocarpaceae) at the higher elevation (ca. 150 m).
Data Deficient given the lack of recent information or collections. Rhodamnia waigeoensis is presently known only from two collections. The specimen on the type label indicates that the species was common locally at the time of its collection nearly sixty years ago. A vegetation type similar to that of the type gathering occurs on the island of Rauki, where the species also may occur. While the reported ethnobotanical use of Rhodamnia waigeoensis for cigarette making may lend the species some protection, it also may have encouraged overexploitation.
Kikir (in the Malayan language).
The herbarium label indicates that the leaves are used for making cigarettes.
Rhodamnia waigeoensis belongs in the “pearly”group of species given its nacreous indumentum (
Rhodamnia waigeoensis differs from Rhodamnia novoguineensis by its thickly coriaceous leaves (vs. thinly coriaceous in Rhodamnia novoguineensis), consistently basal acrodromous leaf venation (vs. even or uneven suprabasal acrodromous in Rhodamnia novoguineensis), dense abaxial laminar indumentum with yellowish trichomes (vs. relatively sparse and whitish in Rhodamnia novoguineensis), solitary flowers (vs. triads or few-flowered racemes in Rhodamnia novoguineensis), thicker (0.5–0.7 mm wide) and rigid pedicels (vs. ca. 0.3 mm thick and flaccid in Rhodamnia novoguineensis).
Waigeo Island is part of the Raja Ampat Islands of Indonesian New Guinea. The region harbors unusual vegetation assemblages (van
The label of the type specimen refers directly to the xerophytic vegetation located behind the small village of Waifoi on the east bank of Majalibit Bay. Takeuchi (2003a, b) reported that the Waigeo ultrabasic vegetation resembles the pioneer communities on the ultrabasics at the Kamilai Wildlife Management Area (KWMA) in the Bowutu Mountains (Morobe Province, Papua New Gueina). Communities at KWMA can be topographically unstable due to landslides, but in general appearance and composition are similar to those on Waigeo. However,
A xerophytic vegetation similar to that occuring on the hills upslope of Waifoi, the village near the type collection, was encountered elsewhere by van Royen (1960: 39, 41) in the Kambelay Hills and the Go Isthmus of Waigeo Island. This general type of xerophytic vegetation is said to recur on Rauki Island, which lies northwest of Kabaré Bay, where it occurs at the higher elevations (probably less than ca. 40 m, but reported by van Royen [p. 45] as 25 m) along the southern end of the island (van
Van Royen (1960: 32) described the soils underlying the xerophytic vegetation on Waigeo as “sandy brown clays with much limestone”. The relatively open vegetation on the slopes was indicated as being spare of trees but conspicuous in its presence of shrubby Myrtaceae. Noted specifically for Myrtaceae (van
West Papua (Papua Barat; as Radjah Ampat on label), Waigeo Island, Waifoi on E bank of Majalibit [= Mayalibit] Bay, 18 Jan 1955, P. van Royen 5227 (L).
Phenology. Flowering September through December; fruiting presumably October through at least February (
Distribution. Northeastern Australia to east-central and southeastern Papua New Guinea;rainforests (including ultrabasics at Kamiali in Papua New Guinea) to margins of anthropogenically derived grassland (Brass 28176); near sea level to 500 m.
Conservation status. Least Concern (IUCN, 2010) based on its known distribution and abundance.
Comments. The known range of Rhodamnia sharpeana is now considerably wider than previously understood (
Specimens examined. Papua New Guinea. Morobe Province, Kamiali Wildlife Management Area; alluvial floodplain along Saia (Sela) River, 7°21.5'S, 147°08'E, ca. 25 m, 25 May 2000, W. Takeuchi 14405(K!); near mouth of Saia River at mount of Hessen Bay, alluvial flatland forest, sea level, 7°21.7'S, 147°08.3'E, common riverbank shrub, 4–5 m height but forest occurrences much taller; leaves dry-textured, adaxially dark dull green, petals white, W. Takeuchi & A. Towati14841(K!); ridge to Blue Mt, 7°17'29"S, 147°05'12"E, ca. 762 m, 28 Feb 2005, W. Takeuchi & D. Ama 18977(BISH (736568! and 736569!), LAE [n.v.]). Milne Bay Province, Sudest [= Tagula or Tagala] Island, (inland from) Rambuso, 300 m, 20 Sep 1956, L. J. Brass 28176(BISH 733105!; A n.v., K n.v., L n.v., US n.v.).
Sixteen new species of Rhodamnia, including the five newly proposed here, have been described since Scott’s (1979) generic revision (
Additional studies still are clearly needed in Rhodamnia. The phylogeny of the genus has never been established, and the hypothesized informal groups of Rhodamnia (
Efforts also should be made to recollect all species of Rhodamnia in New Guinea given the paucity of specimens for many species and the present inability to assign conservation threat assessments with high levels of confidence. Two suggestions for collecting in remote, biodiverse areas are worthy of repeating. First, as
I thank the curators at A, BM, BISH, BRI, CANB, K, L, MEL, MO, NSW, NY, QRS, PH, RSA, and US for loaning material critical to this project. The Herbarium of the Arnold Arboretum (A) kindly gave permission to capture digital images of Brass 22718and van Royen 5556. Krista Anandakuttan provided the illustrations of Rhodamnia makumak and Rhodamnia toratot while in residency at the Bishop Museum. Drs. Michael Kiehn (W) and William A. Weber (COLO) translated passages from German. Support for curatorial work of New Guinea specimens was supported in part by the National Science Foundation (DEB 1057453) to Dr. Shelley James (BISH), whom I thank for forwarding digital images. Casey Currier also assisted with the preparation of some digital figures. Dr. Kanchi Gandhi (A) checked the derivation of Latin epithets; and Dr. Peter Wilson (NSW) and Bryan Simon (BRI) contributed to and confirmed some observations of Rhodamnia toratot and Rhodamnia makumak. I thank Dr. Wayne Takeuchi and an anonymous reviewer for their suggestions. This paper represents Contribution no. 2012-015 to the Pacific Biological Survey.