





(C) 2012 Lubbert Y.T. Westra. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The taxonomic revision of the infrequently collected genus Tetrameranthus by
Annonaceae, taxonomy, Neotropics, Tetrameranthus
An as-yet unknown species of the rare genus Tetrameranthus has been collected a few times in French Guiana and also once in neighboring Amapá, Brazil. The material was too incomplete to justify publication. Recently a new collection from French Guiana came in bearing ripe fruits and also reported to have a white flower. This made us decide now to formally publish this new species. We then also decided to update the revision of the genus Tetrameranthus (Westra, 1985), with the inclusion of Tetrameranthus globuliferus published later (Westra, 1988), adding the present new species as well. Some, mostly color, photographs of this seldom collected genus serve as an illustration.
Taxonomic historyThe Annonaceae form a large pantropical family, the largest family in the order Magnoliales, currently with (about) 129 genera and >2000 species (
The Neotropical genus Tetrameranthus was described by
Over fifteen years later Fries added a second species, Tetrameranthus macrocarpus R.E.Fr., from Southeastern Colombia (
Four more species were added in later years, viz. Tetrameranthus laomae D.R.Simpson from Eastern Peru (
Based on recent molecular data it was shown that Tetrameranthus belongs to an early branching lineage within Annonaceae (
The generic name is composed of the Ancient Greek word elements “tetra” (four), “meros” (part), and “anthos” (flower), referring to the four-parted flowers.
Morphology Vegetative partTetrameranthus isa genus of shrubs to (large) trees. As for all Annonaceae, the leaves are simple, entire, pinnately veined, symmetrical, and lack stipules, generally of moderate to fairly large size (greatest length ±30 cm). In contrast to the rest of Annonaceae, where the leaves are in two alternate rows almost without exception (
This rosette-like appearance of leaf disposition is suggestive of a number of other plant families such as Sapotaceae, but not of Annonaceae. This could explain in part why Tetrameranthus might seem rare, i.e., simply because many botanists do not identify it as Annonaceae.
lnflorescencesInflorescences in Tetrameranthus are truly axillary. As for the structure of the inflorescence in Annonaceae in general, the reader is referred to publications by
For these reasons the basal part of the first-order flower stalk up to the articulation and including the whorl of bracts is referred to as peduncle, while the part above the articulation is termed pedicel. The stalks of second-order flowers, then, are termed pedicels in their entirety.
FlowersAs in most Annonaceae, the perianth consists of one whorl of sepals and two whorls of petals (Figure 1A). The whorls are 4-merous in at least five of the seven species, although incidentally a single deviating flower might occur, e.g. a 3-merous flower on one specimen of Tetrameranthus laomae, and a 5-merous flower on the type collection of Tetrameranthus pachycarpus. Such phenomena are not unusual throughout flowering plants in general, and have been observed in other Annonaceae genera (
The sepals are small in relation to the petals. They are free or connate just at the base. The aestivation is imbricate, only observable in very young buds as the sepals soon spread. The sepals drop after flowering.
The petals vary from rather fleshy (e.g. Tetrameranthus duckei, Figure 2C) to rather thin (Tetrameranthus laomae, Tetrameranthus umbellatus, Figure 2F), those of the outer whorl being somewhat larger or broader than those of the inner whorl. The photographs also showcurved petals in Tetrameranthus duckei in the living condition, while in Tetrameranthus umbellatus (and possibly also in Tetrameranthus laomae) the petals appear rather flat at least before anthesis. The petals are adorned with a callus or callus-like tissue at the base on the inner side: this is an area of varying size, depending upon the species, which is devoid of indument. In Tetrameranthus duckei the callus appears as a protruding hump (Figure 2C), particularly on the inner petals where it is even larger than on the outer petals. In the other species the callus is smaller in relation to the size of the whole petal than in Tetrameranthus duckei.
The convex torus bears mostly numerous stamens and a mostly rather small number of free carpels in the center. The stamens have a short filamental part, a thick connective capped by a massive, more or less conical or flat shield, and an extrorse to latrorse anther. The carpels contain two (exceptionally three), lateral, superposed ovules, and have on top a sessile stigma which varies from trilobed to an irregularly lobed disc.
Curculionidae beetles have been observed as pollinators in Tetrameranthus duckei (
Depending on the species, the number of free monocarps varies from 1–15. The ellipsoid to oblongoid monocarps are fleshly, indehiscent, and two-seeded or, due to abortion of one ovule, one-seeded (rarely three seeds develop) (Figure 1C–G). The seeds are laterally attached, with the lowest one near the base, and are ascending, thus resulting in the characteristic oblique constriction seen in monocarps with >1 seed on herbarium specimens. In fresh fruits the constriction is less obvious, and in very thick-walled monocarps of some of the species (e.g. Tetrameranthus guianensis, Tetrameranthus pachycarpus) it becomes practically indistinguishable. In herbarium material the fruit wall is smooth in most species, but becomes shriveled in Tetrameranthus globuliferus (
The seeds are quite large in relation to the fruit body; they are slightly compressed dorsiventrally, and possess ruminations in the shape of fairly numerous lamellae protruding from the seed coat into the interior almost to the middle.
IndumentThe indument of Tetrameranthus consists of stellate hairs with 2–10 rays, varying with the species. In addition to stellate hairs, simple hairs are present in varying densities, also depending upon the species. Most genera of Neotropical Annonaceae have simple hairs, or have stellate hairs beside simple hairs in a small percentage of the species only (e.g. Annona including Rollinia). The notable exception (apart from Tetrameranthus) is Duguetia: in this genus most species even have scales, rather than stellate hairs (
Indument in Tetrameranthus isfound especially on young vegetative parts and inflorescences. Some of it persists on the primary vein of the leaves, and to a lesser extent on the secondary veins, mainly on the lower side; it is also seen on petioles and branchlets in the leafy zone. In Tetrameranthus umbellatus stellate hairs are also found diffusely spread over the leaf surface, especially on the abaxial side. In Tetrameranthus laomae, too, scattered stellate hairs may be spotted on the lower leaf surface.
Floral parts, with the exception of stamens and the callus area on petals mentioned earlier, are usually covered with a dense indument of stellate hairs. Carpels, when enlarging into monocarps, quickly become glabrous.
The trichome length on vegetative parts (except in a very young stage, and persisting near axils) does not exceed 0.1–0.2 mm in Tetrameranthus laomae and Tetrameranthus umbellatus: these two species thereby are easily distinguished from the other ones, where considerably longer (to 0.5 mm, or even more) and stiffer trichomes are found, next to simple hairs of the same size. Trichomes on inflorescences and flower parts may reach a somewhat larger average size than those on vegetative parts (this we did not investigate in detail).
Taxonomic treatmenthttp://species-id.net/wiki/Tetrameranthus
Tetrameranthus duckei R.E.Fr.
Trees or shrubs. Leafy twigs and most floral parts sparsely to densely covered with stellate to simple hairs to glabrous. Leaves spirally arranged, often concentrated towards the end of the branches, primary vein impressed to slightly raised on the upper side. Inflorescences axillary, 1-flowered to several-flowered and umbel-like, bracts up to 4 below the articulation. Flowers bisexual, 4-merous or less often 5–6-merous, white to yellow or cream; sepals 4(–6), imbricate, free or basally connate; petals 8(–12), free, subequal, imbricate, much longer than the sepals, often with a callus at the inner base; stamens numerous, connective shield discoid, either flat, cushion-shaped, or with a conical prolongation; carpels c. 5–30, ovules 1–2(–3), lateral, stigma sessile, more or less lobed. Fruit apocarpous; monocarps 1–15, free, sessile or sometimes narrowed into a short and thick stipe-like base, indehiscent, sometimes constricted, wall rather thick (1–7 mm) and fleshy. Seeds 1–2(–3) per monocarp, lower one near the base, upper one(s) lateral.
Seven species in the Amazon regions of Venezuela, Colombia, Brazil, Peru, and Ecuador, but also in the Colombian state of Chocó and in French Guiana and neighboring Amapá, Brazil.
| la | Young plant parts covered with stellate hairs ≤0.2 mm long; primary vein mostly flat (or slightly raised or slightly impressed) above | 2 |
| 1b | Young plant parts covered with stellate and simple hairs ≥0.5 mm long; primary vein impressed (exceptionally almost flat) above | 3 |
| 2a | Inflorescences with up to 5 umbellately arranged flowers; peduncle manifest, ≥5 mm long (Amazonian Peru and Brazil) | Tetrameranthus umbellatus |
| 2b | Inflorescences 1-flowered; peduncle inconspicuous, <1 mm long. (Amazonian Peru and Brazil and the Colombian states of Amazonas and Chocó) | Tetrameranthus laomae |
| 3a | Monocarps globose or almost; perianth (as far as known) of 6-merous whorls (Amazonian Ecuador) | Tetrameranthus globuliferus |
| 3b | Monocarps ellipsoid, oblongoid or fusiform; perianth of 4-merous or less often 5-merous whorls | 4 |
| 4a | Monocarps, both 1- and more-seeded ones, ellipsoid or fusiform, without constriction or with a weak constriction | 5 |
| 4b | 2-Seeded monocarps oblongoid, with a manifest oblique constriction about the middle | 6 |
| 5a | Monocarps 7–15, 35–60 by 20–30 mm, wall shriveled in sicco; petioles ≤10 mm long (French Guiana and the Brazilian state of Amapá) | Tetrameranthus guianensis |
| 5b | Monocarps 1–3, to c. 70 by 40 mm, wall not shriveled in sicco; petioles ≥20 mm long (Amazonian Peru, vicinity of Iquitos) | Tetrameranthus pachycarpus |
| 6a | Monocarps ≥35 mm in diam.; tall tree (Amazonian Colombia) | Tetrameranthus macrocarpus |
| 6b | Monocarps ≤25 mm in diam.; shrub or small tree ≤12 m (Amazonian Colombia, Venezuela, and Brazil) | Tetrameranthus duckei |
http://species-id.net/wiki/Tetrameranthus_duckei
Figs 1C, D, 2A–D; Map 1Ducke RB 23919 (holotype S; isotypes RB, S), Brazil, Amazonas: Manaus, Estrada do Aleixo, km 7, 14 June 1933.
Shrub or tree, 3–12 m tall, 4–8 cm diam., young twigs and petioles densely to rather densely covered with brown, stellate hairs >0.5 mm long, becoming glabrous. Leaves: petioles 10–40 mm long, 1.5–4 mm diam.; lamina narrowly elliptic to narrowly obovate, 10–25 by 3–10 cm (index 2.7–4), chartaceous to coriaceous, dull or slightly shiny brown or greenish brown above, dull brown or greenish brown below in sicco, rather densely covered with stellate hairs on primary vein, otherwise glabrous above, rather densely to sparsely covered with stellate hairs on primary vein and secondary veins, otherwise mostly glabrous below, the stellate hairs similar to those on branchlets, base acute, apex acuminate (acumen 5–25 mm long), primary vein impressed above, secondary veins 8–12 on either side of primary vein, impressed above, loop-forming, shortest distance between loops and margin 1.5–5 mm, or not loop-forming in basal part, tertiary veins slightly raised, flat, or indistinct above, percurrent to reticulate. Inflorescences 1(–2)-flowered, peduncles 5–15 mm long, c. 1.5 mm diam., fruiting peduncles to c. 3 mm diam., bracts 4, narrowly triangular, 3–6 mm long, soon falling after flowering, pedicels 10–25 mm long, c. 1.5 mm diam., fruiting pedicels to c. 40 mm long, 3 mm diam., peduncles and pedicels densely covered with stellate hairs, becoming glabrous. Flowers green, turning yellow in vivo; sepals elliptic to obovate, free, 5–7 mm long, outer side densely covered with stellate hairs; outer petals ovate, 20–25 by 9–12 mm, inner base with fleshy and longitudinally grooved callus 5–6 mm long and extending across the whole width, inner petals narrowly ovate to ovate, 15–22 by 6–9 mm, with similar callus to c. 8 mm long, outer side of petals densely covered with stellate hairs, the callus on the inner side glabrous; stamens 2–2.5 mm long, connective shield conical or acuminate, 1–1.5 mm long, more or less curved toward the center. Monocarps 1–6, green or shiny green, turning green-yellow in vivo, brown to dark brown in sicco, ellipsoid or oblongoid to narrowly so, 25–65 by c. 20(–25) mm, with (2-seeded forms) or without oblique constriction, apex a thick obtuse beak 2.5–10 mm long. Seeds 1–2 per monocarp, to c. 35 by c. 20 mm.
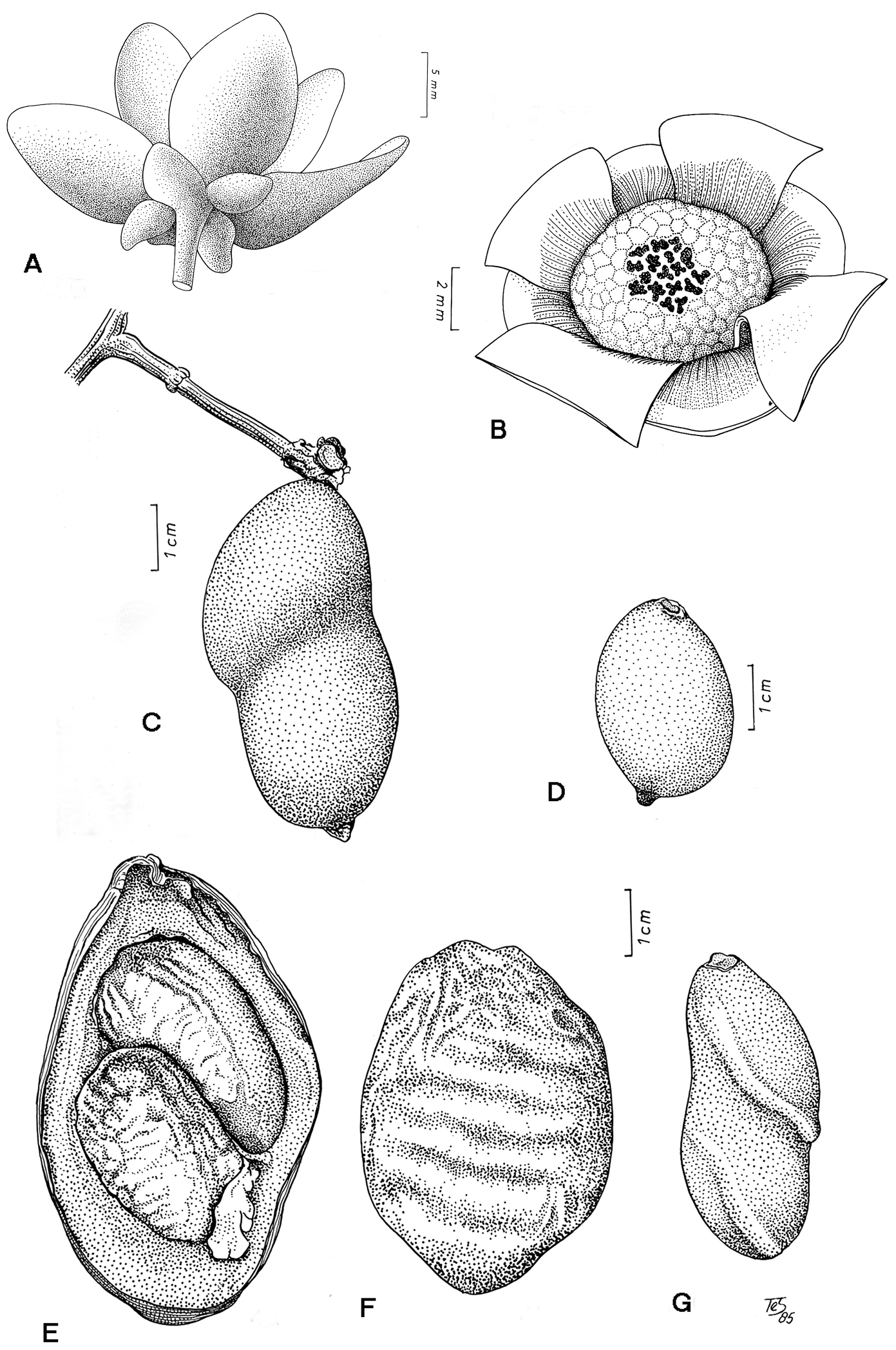
Tetrameranthus umbellatus Westra A Young flower (Tunquí 62, U) B Androecium and gynoecium of same. Tetrameranthus duckei R. E. Fr. C Two-seeded monocarp (Rodrigues & Coêlho 3835, U) D One-seeded monocarp (Morawetz et al. 21-9883, U). Tetrameranthus pachycarpus Westra E, F Two-seeded monocarp cut open and seen from outside (Foster 4271, NY). Tetrameranthus umbellatus Westra G Two-seeded monocarp (Huashikat 613, U).

Tetrameranthus duckei R. E. Fr. A Two-flowered inflorescence B Flowers at early anthesis C Flower at late anthesis, liberation of pollen (Webber, 1981) D Fruit. Tetrameranthus laomae D.R.Simpson E Ripe fruit, also sectioned to show seeds. Note articulation at base of fruiting stalk. Tetrameranthus umbellatus Westra F Flowers G Young inflorescence, detail of herbarium specimen (Morawetz & Wallnöfer 14-81085, U). A, E–F Photos by W. Morawetz, B–D Photos by A. C. Webber.
Amazonian regions of Venezuela (Amazonas), Colombia (Guainía), and Brazil (Amazonas, most common in Manaus and vicinity).
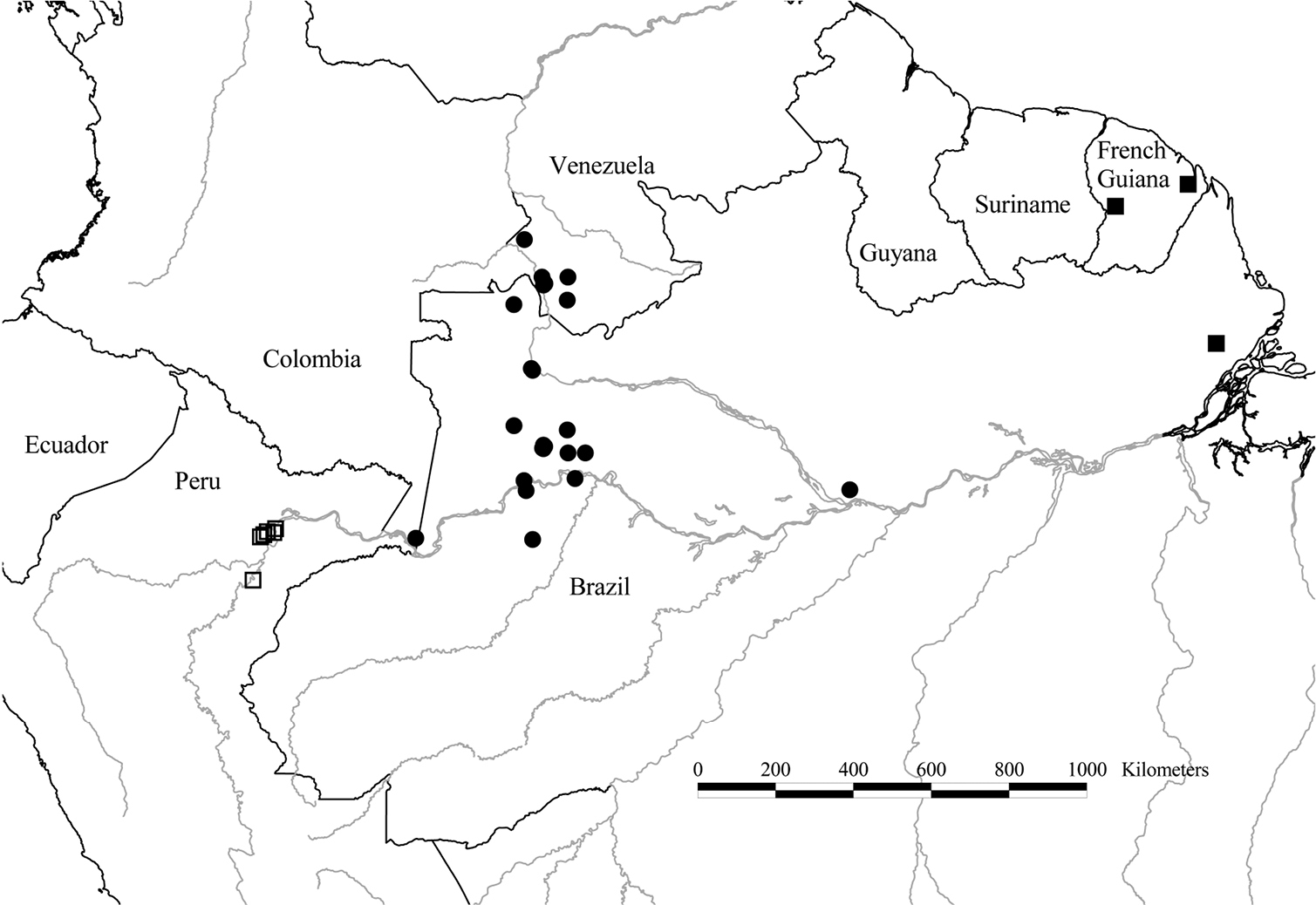
Distribution of Tetrameranthus duckei R. E. Fr. (●), Tetrameranthus guianensis Westra & Maas (■), and Tetrameranthus pachycarpus Westra (□).
Mostly in low forest or shrub vegetation (campina, campinarana, Amazonian caatinga, and bana) on white sand. At low elevations up to c. 200 m. Flowering and fruiting: throughout the year.
Colombia. Guainía: Puerto Colombia (opposite Venezuelan town of Maroa) and vicinity, alt. 800–850 ft, Schultes et al. 18157 (US).Venezuela. Amazonas: Mun. Guainía, along road from Maroa to Yavita, Acevedo-R. et al. 10250 (U); Lower Río Guainía, Raudal Lombriz, 2 km from mouth of Río Casiquiare, 140 m, Aymard et al. 9751 (MO, U); Río Casiquiare, below Capibara (“Capihuara”), Colella et al. 1879 (U); Río Casiquiare, El Porvenir, Colella et al. 2170 (U); San Carlos de Río Negro, Christenson 1386 (US), Liesner 6744 (MO, U), 7590 (MO, U), 8598 (MO, U), 8838 (MO, U), Steyermark & Bunting 102721 (NY, US); Río Negro, base of Piedra de Cucuy, 100–200 m, Maas et al. 6879 (INPA, NY, U); Río Pasimoni, 80 m, Velazco 1954 (MO, U). Brazil. Amazonas: Manaus and vicinity, Almeida INPA 3580 (INPA), L. Coêlho INPA 3673 (INPA, S), Ducke 1908 (F, NY), Ducke RB 35313 (RB, S), Ferreira 79/57 (S), Miralha et al. 230 (INPA, U), Morawetz et al. 21-9883 (WU), 21-23883 (WU), 22-19883 (WU), 24-12983 (WU), Personel of Centro de Pesquisas Florestais INPA 6232 (INPA, S), Plowman et al. 12647 (U), Prance et al. 2721 (INPA, NY, US), 3816 (NY, US), 4679 (NY, US), Rodrigues & L. Coêlho 2937 (U), Rodrigues & Almeida 3068-A (U), Rodrigues & Lima 3454 (U), Rodrigues & D. Coêlho 3839 (INPA, U), Rodrigues 8742 (INPA), Webber 162 (U), 163 (U); Reserva Florestal Ducke, Igarapé Acará, Ribeiro et al. 1501 (INPA), 1749 (INPA, U), Sothers et al. 757 (INPA, U); Rio Negro, at its confluence with Rio Vaupés, Serra Canaleão, 150 m, Stevenson et al. 1002 (NY, U); Rio Javari, behind Estirão de Equador, Lleras et al. P17302 (NY, U); mouth of Rio Vaupés, Pires et al. 7473 (S).
Venezuela: Banayo (Liesner 7590), Cuchara (Liesner 6744), Majagua (Velazco 1954), Palo de cuchara (Liesner 7590). Brazil: Envira (Ferreira 79/57).
In a previous paper (Westra, 1985) there was some doubt about the identity of the collection Lleras et al. P17302. It should be regarded as no more than an extreme form of Tetrameranthus duckei, with pedicels to c. 40 mm long and outer petals to c. 15 mm wide.
http://species-id.net/wiki/Tetrameranthus_globuliferus
Fig. 3D–G; Map 2Lawesson et al. SEF 8779 (holotype AAU; isotypes AAU, QCA, QCNE, U), Ecuador, Orellana: Añangu, Parque Nacional Yasuní, 260–350 m, May–June 1986.
Medium-sized tree, >10 cm diam., young twigs and petioles densely covered with pale brown, stellate hairs >0.5 mm long. Leaves: petioles 4–8 mm long, 4–6 mm diam., lamina narrowly obovate, 27–37 by 9–15 cm (index 2.8–2.9), chartaceous, shiny green above in vivo, greenish brown above, pale greenish brown to brown below in sicco, rather densely covered with stellate hairs >0.5 mm long on primary vein, to rather sparsely so on smaller veins on both sides, base acute to attenuate, to obtuse or rounded at the extreme base, apex acute to acuminate (acumen to c. 10 mm long), primary vein flat to slightly raised above, secondary veins 20–25 on either side of primary vein, flat to impressed above, loop-forming, shortest distance between loops and margin 1.5–3 mm, or not loop-forming, tertiary veins flat to raised above, percurrent to more or less reticulate. Inflorescences 1-flowered; peduncles 3–5 mm long, 3–4 mm diam., fruiting peduncle c. 5 mm diam., bracts [4?] narrowly oblong or narrowly triangular, 4–5 mm long, falling after flowering, pedicels {18–30} mm long, {4–6} mm diam., densely covered with brownish, stellate hairs; flowers with perianth in 6-merous whorls, cream with the inner petals yellow at the inner base in vivo; sepals broadly ovate-triangular, connate at the very base, {6–11} by {5–10} mm; outer petals elliptic to ovate, {30–45} mm long, {10–25} mm wide, with small callus at the inner base; inner petals {30–40} mm long, {5–10} mm wide, more or less narrowed toward the base, with larger callus, outer side of petals densely covered with stellate hairs, the callus on the inner side glabrous; stamens {2–2.5} mm long, connective shield flat, cushion-shaped. Monocarps 2–7, globose or almost, green in vivo, brown in sicco, c. 40 mm diam., wall strongly shriveled in sicco. Seeds 1–2 per monocarp, 25–30 by 15–20 mm.

Tetrameranthus guianensis Westra & Maas A Single-flowered inflorescence after shedding of perianth and stamens B A twig C Ripe fruit, also sectioned to show seed. Tetrameranthus globuliferus Westra D, E Twig with inflorescences (Pérez C. & Santillán 4404, QCA) F, G Flower seen from the side and from above (same). A–C Photos by D. Sabatier, D–G Photos by A. J. Pérez C.
Ecuador (Orellana). Only known so far from Parque Nacional Yasuní.
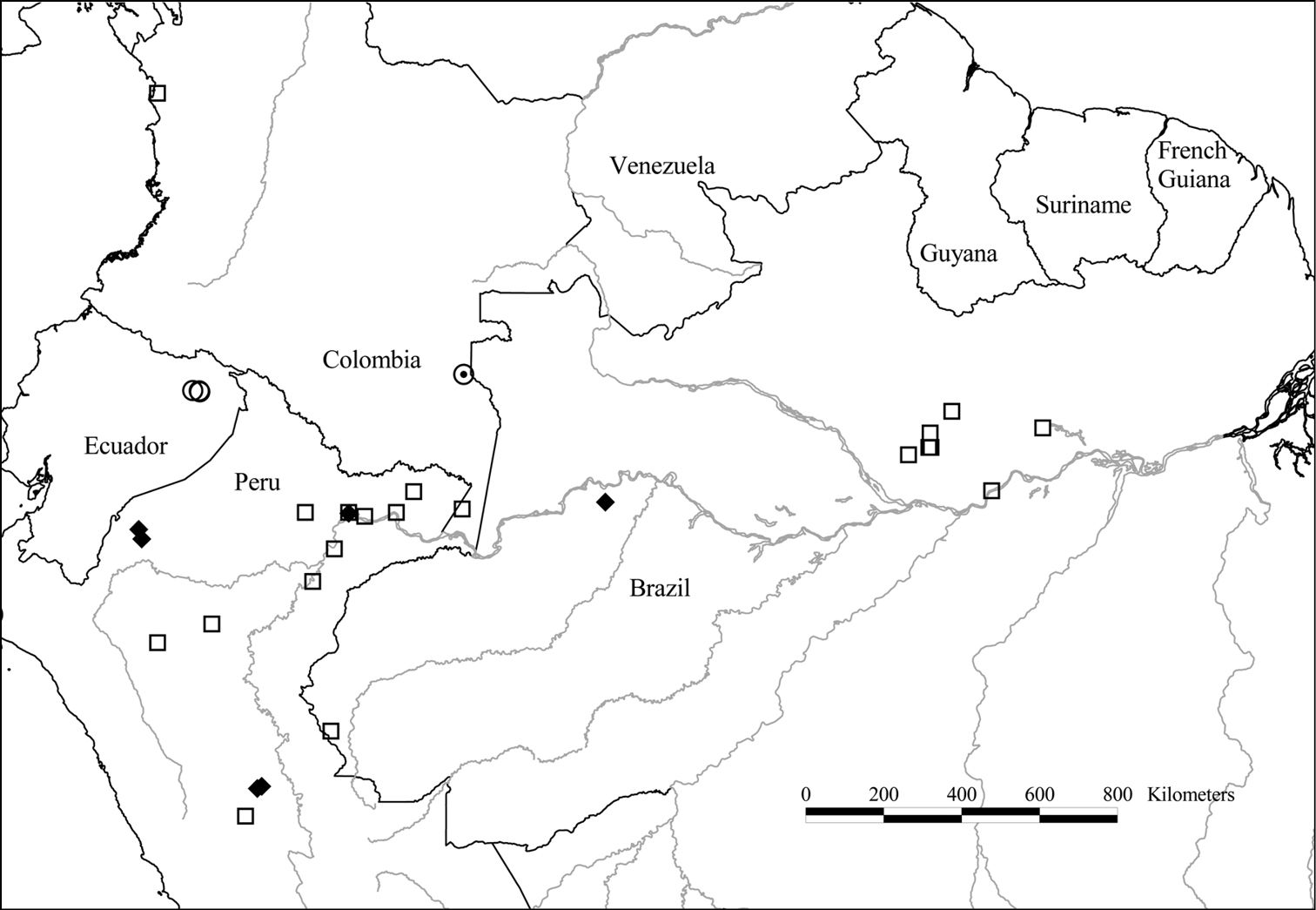
Distribution of Tetrameranthus globuliferus Westra (○), Tetrameranthus laomae D. R. Simpson (□), Tetrameranthus macrocarpus R. E. Fr. (◉), and Tetrameranthus umbellatus Westra (◆).
In rain forest on terra firme. At elevations of 200–400 m. Flowering recorded in November; fruiting recorded in May, June, August.
Ecuador. Orellana: NW corner of Parque Nacional Yasuní, 355–365 m, Korning & Thomsen 47626 (AAU, U); Parque Nacional Yasuní, E of Juan Tapuy’s finca, 250 m, Pitman & Delinks 1440 (MO, QCA, U).
The description of the flower was largely made from a very recent photograph of a freshly collected twig of Pérez & Santillán 4404 (QCA). Measurements between { } were made on two flowers preserved in alcohol kept in QCA and were kindly supplied to us by Álvaro J. Pérez C.
Á. J. Pérez also reports to us a clustered occurrence of Tetrameranthus globuliferus: he found few individuals all close to the one that he collected, but did not spot any more around the trails of the Yasuní Scientific Station.
The species has been listed as near-threatened on the IUCN red list (
urn:lsid:ipni.org:names:77118748-1
http://species-id.net/wiki/Tetrameranthus_guianensis
Fig. 3A–C; Map 1A speciebus ceteris hujus generis pilis stellatis vel simplicibus validis et monocarpiis rugosis in statu sicco distinctus.
Sabatier & Gonzalez 5387 (holotype CAY; isotype L), French Guiana, Savane-roche Virginie, Parcelle SRV2, 4°11'N, 52°9'W, 18 March 2008.
Tree, 8–25 m tall, 10–25 cm diam., slash yellow-orange (Mori et al. 23521); young twigs and petioles densely covered with brownish, stellate and simple hairs >0.5 mm long. Leaves:petioles 5–10 mm long, 3–4 mm diam., more or less thickened toward the base; lamina narrowly obovate to obovate-elliptic, 14–26 by 5–10 cm (index 2.4–2.9), chartaceous, shiny green above in vivo, dark brown above and pale brown below in sicco, rather densely covered mainly on large veins to rather sparsely covered with stellate hairs >0.5 mm long or glabrous elsewhere above, densely covered with stellate hairs on large veins, rather densely to sparsely so or almost glabrous elsewhere below, base acute to attenuate, apex abruptly acuminate (acumen 2–12 mm long), primary vein impressed above, secondary veins 14–19 on either side of primary vein, impressed above, loop-forming, shortest distance between loops and margin 1–3 mm, or not loop-forming, tertiary veins impressed above, percurrent to reticulate. Inflorescences 1-flowered, only seen in postfloral and fruiting stages, peduncles c. 10 mm long, c. 3 mm diam., fruiting peduncles to c. 5 mm diam., pedicels c. 20 mm long, c. 3 mm diam., fruiting pedicels to c. 5 mm diam., peduncles and pedicels densely covered with brownish, stellate and simple hairs; sepals not seen; petals whitish (fide collectore) in vivo, estimated to be c. 35 by 40 mm; stamens not seen. Monocarps 7–15, ellipsoid to fusiform, yellowish green to yellowish orange in vivo, pale brown in sicco, 35–60 by 20–30 mm, apex obtuse, rounded, or bluntly pointed, with or without weak oblique constriction, wall shriveled in sicco. Seeds 1–3 per monocarp, 25–30 by 15–20 by 15 mm.
French Guiana and the adjacent Brazilian state of Amapá.
In forests. At an elevation of c. 100 m. Flowering recorded in December; fruiting recorded in March, July, and December.
Tetrameranthus guianensis is the first species of the genus reported from the Guianas. It is distinct from other species of Tetrameranthus by a dense cover of coarse stellate and simple hairs on all vegetative parts. Like the Ecuadorian Tetrameranthus globuliferus, it has shriveled fruit walls in dry condition.
As no complete flowers were available in herbarium material, description of floral characters is based in part on field observations of a single living flower at distance. The collector, D. Sabatier, has informed us that an attempt to collect that flower which was high up in a large tree and out of reach had failed. Sabatier (pers. comm.) notices 5 scars per whorl on the receptacle (compare Figure 3A) implying that we have a 5-merous flower here! This definitely requires confirmation from further collections, though.
Two sterile collections, namely Mori et al. 23521, 23674, also from French Guiana, seem to come quite near this species, differing mainly in the leaf shape (obovate-elliptic, rather than narrowly obovate), and the less dense and more coarse indument of stellate hairs of comparable size. It concerns trees of 8 m, 10 cm diam., and 15 m, 12 cm diam., respectively, from non-flooded moist forest. More material, and more complete in particular, is needed here.
French Guiana. Sinnamary River, above Petit Saut, between Crique Plomb and Crique Tigre, 500 m above Saut Tigre in area to be inundated by waters of Petit Saut Dam, Mori et al. 23521 (CAY, NY, U), Mori et al. 23674 (CAY, NY, U); Rivière Grand Inini, Basin of Maroni River, Arbre II-59, 3°40'N, 53°50'W, Sabatier & Prévost 3084 (CAY, P, U); Savane-roche Virginie, Parcelle SRV2, 4°11'N, 52°9'W, Sabatier 5784 (CAY, WAG). Brazil. Amapá: Rio Araguari, upland plant, Pires et al. 51490 (MG).
http://species-id.net/wiki/Tetrameranthus_laomae
Fig. 2D, E; Map 2Soria S. 64 (holotype F), Peru, Loreto: Alto Amazonas, Distr. Yurimaguas, road from Yurimaguas to Tarapoto, km 19 from Yurimaguas, 115 m, 11 March 1969.
Tree, 7–35 m tall, 11–45 cm diam., young twigs and petioles densely covered with whitish, stellate hairs <0.2 mm long, becoming glabrous. Leaves: petioles 10–20 mm long, 1–1.5 mm diam.; lamina obovate to narrowly obovate or narrowly elliptic-obovate, 6–24 by 2–7 cm (index 2.4–3.7), chartaceous to thinly coriaceous, shiny to dull greenish brown above, dull greenish brown below in sicco, rather densely to sparsely covered with stellate hairs <0.2 mm long on primary vein, otherwise sparsely so to glabrous on both sides, base attenuate, decurrent along petiole, apex acute or acuminate (acumen (0–)7–20 mm long), primary vein raised to almost flat above, secondary veins 7–10 on either side of primary vein, more or less raised above, loop-forming, shortest distance between loops and margin 2–4 mm, or not loop-forming in basal part, tertiary veins raised above, reticulate and often tending to form intersecondaries. Inflorescences 1-flowered, peduncles 0–1 mm long, bracts 3, narrowly triangular or narrowly oblong, 1–2.5 mm long, soon falling after flowering, pedicels 7–15 mm long, 1–1.5 mm diam., fruiting pedicels to c. 20 mm long, 2.5–4 mm diam., densely to rather densely covered with stellate hairs, becoming glabrous. Flowers green to yellowish, green with yellow center, or yellow in vivo; sepals broadly elliptic to broadly ovate, 2.5–3 by 2–2.5 mm, outer side densely covered with stellate hairs; outer petals ovate-elliptic, 10–16 by 5–8 mm, inner petals narrowly elliptic, 8–11 by 3–4 mm, outer side of petals rather densely covered with stellate hairs, the inner base glabrous; stamens c. 1 mm long, connective shield cushion-shaped, flat. Monocarps 4–13, green, maturing orange or red in vivo, black in sicco, ellipsoid to oblongoid, 2-seeded forms 30–45 by 15–<20 mm and with oblique constriction, 1-seeded forms smaller and without constriction, apex mostly rounded. Seeds 1–2 per monocarp, to c. 25 by 20 mm.
Amazonian Peru (Loreto, Pasco) and Brazil (Acre, Amazonas, Pará), and the Colombian states of Amazonas and Chocó.
In forest on terra firme, on clay or sandy soil. At low elevations to c. 350 m. Flowering and fruiting: throughout most of the year.
Colombia. Amazonas: Mun. Leticia, Corregimiento de Tarapacá, Parque Nacional Natural Amacayacu, Cabaña Pamaté, 100 m, Rudas et al. 2611 (MO, U). Chocó: Parque Nacional de Utría, 0–100 m, García C. & Agualimpia 398 (MO, U). Brazil. Acre: vicinity of Serra da Moa, Prance et al. 12236 (MO, NY, U). Amazonas: Mun. Itapiranga, Rio Uatumã, above confluence of Rio Uatumã with Rio Pitinga, Cid et al. 824 (MO, NY, U); Mun. Presidente Figueiredo, near Represa de Balbina, Cid et al. 6657 (NY, U), 8084 (NY, U), Thomas et al. 5271 (NY, U), 5358 (NY, U). Pará: Mun. Oriximiná, Rio Trombetas, Lago Maincoé, 8 km NE of Mineração Santo Patricia, 80 m, Martinelli 7338 (NY, U). Peru. Loreto: Guarnición Pijuayal, near Pebas, 130 m, Díaz S. et al. 571 (MO); Distr. Las Amazonas, “Roca Eterna”, 120–130 m, Grández & Jaramillo 2850 (MO, U); Distr. Las Amazonas, Explornapo Camp, near Sucusari, 100–140 m, Pipoly et al. 13386 (MO, U), 13407 (MO, U), 13496 (MO, U), 14575 (MO, U); Prov. Mariscal Ramón Castilla, Upper Río Yaguas, tributary of Río Putumayo, 80 km NE of Pebas, 140 m, Ríos et al. 464 (U); Prov. Ucayali, Sapuena, Jenaro Herrera, 130 m, Vásquez et al. 12012 (MO, U); Prov. Maynas, Sargento Lores, Esperanza, 120 m, Vásquez & Jaramillo 13249 (MO, U). Pasco: Prov. Oxapampa, Cabeza de Mono, Río Iscozacín, Palacazu Valley, 320 m, Gentry et al. 41743 (MO, U); Prov. Oxapampa, Distr. Iscozacín, Pariona & Ruíz C. 1038 (MO).
Peru: Sacha anona (Pariona & Ruíz C. 1038).
Peru: used for construction work on the countryside (Soria S. 64).
http://species-id.net/wiki/Tetrameranthus_macrocarpus
Map 2Schultes & Cabrera 17091 (holotype S; isotypes COL, GH, U), Colombia, Vaupés: Río Piraparaná, Raudal Coro (“Koro”), 30 August 1952.
Tall tree, young twigs and petioles densely to rather densely covered with stellate hairs >0.5 mm long. Leaves: petioles 20–30 mm long, 3–4 mm diam., slightly thickened toward the base; lamina elliptic to obovate to narrowly so, 14–20 by 5–8 cm (index 2–2.9), coriaceous, glabrous above, rather densely to sparsely covered with stellate hairs on large veins, otherwise glabrous below, base acute to attenuate, apex acuminate (acumen 5–20 m long), primary vein impressed above, secondary veins 9–11 on either side of primary vein, impressed above, loop-forming, shortest distance between loops and margin 1–2 mm, or not loop-forming, tertiary veins flat and inconspicuous above, percurrent to reticulate. Inflorescences 1-flowered; peduncles c. 5 mm long, 2–2.5 mm diam., fruiting peduncles to c. 5 mm diam.; bracts 4, triangular to narrowly triangular, 2–3 mm long, outer side densely covered with stellate hairs, falling after flowering; pedicels 25–30 mm long, c. 2 mm diam., fruiting pedicels to c. 35 mm long, peduncles and pedicels densely covered with stellate hairs, becoming glabrous. Flowers: color not mentioned; sepals broadly triangular-ovate, connate just at the base, 3–4 mm long, outer side densely covered with stellate hairs; outer petals elliptic, 25–30 mm long, 12–15 mm wide, base attenuate into a more or less distinct claw, apex acute to obtuse, callus area at inner base to c. 5 mm long, inner petals similar to outer petals, 20–25 mm long, outer side of petals densely covered with stellate hairs; stamens c. 3 mm long, connective shield with conical prolongation (fide
Colombia (Vaupés). Only known from the type collection.
Not recorded.
This single collection of Tetrameranthus macrocarpusseems to come quite near some forms of the variable Tetrameranthus duckei. It is distinct from the latter primarily by the larger fruit. Moreover, it is reported to be a tall tree (without more precise indication of size), whereas reports for Tetrameranthus duckei indicate a medium-sized tree to about 12 m tall so far. Nevertheless one might wonder if Tetrameranthus macrocarpus is merely an extreme form of Tetrameranthus duckei, but more extensive collecting in the western Amazon region is needed before this question could be answered.
http://species-id.net/wiki/Tetrameranthus_pachycarpus
Fig. 1E, F; Map 1Klug 1216 (holotype NY; isotypes F, U), Peru, Loreto: Mishuyacu, near Iquitos, 100 m, April 1930.
Tree, 4–26 m tall, young twigs and petioles densely to rather densely covered with brown, stellate hairs >0.5 mm long, becoming glabrous. Leaves: petioles 20–30 mm long, 2–4 mm diam., slightly thickened toward the base, lamina elliptic, narrowly elliptic or narrowly obovate, 17–22 by 5–10 cm (index 2.1–3.4), coriaceous, brown in sicco, glabrous except for primary vein above, rather densely to sparsely covered with stellate hairs on large veins and otherwise sparsely covered with stellate hairs to glabrous below, base acute to attenuate, apex obtuse, acute, or acuminate (acumen 1–5 (rarely more) mm long), primary vein impressed to almost flat above, secondary veins 10–15 on either side of primary vein, not loop-forming, or less often loop-forming, shortest distance between loops and margin 2–3 mm, tertiary veins flat and inconspicuous above, percurrent to more or less reticulate. Inflorescences 1-flowered; peduncles 4–5 mm long, 1–2 mm diam., fruiting peduncles to c. 5 mm diam., bracts 2(–more?), narrowly triangular to linear-triangular, 3–5 mm long, outer side densely covered with stellate hairs, falling at or after flowering, pedicels 10–15 mm long, 1.5–2 mm diam., fruiting pedicels to c. 25 mm long, 3–4 mm diam., peduncles and pedicels densely to rather densely covered with stellate hairs, becoming glabrous; flowers yellow in vivo; sepals broadly elliptic, 3–4 mm long, connate at the very base, outer side densely covered with stellate hairs; outer petals narrowly elliptic to oblong, to c. 35 by 16 mm, outer side densely to rather densely covered with stellate hairs, the inner base glabrous, to c. 3 mm long, inner petals narrowly elliptic to oblong, somewhat smaller than outer petals and with slightly larger glabrous inner base; stamens c. 2 mm long, connective shield more or less conical and curved toward the center. Monocarps 1–3, yellow in vivo, brown in sicco, ellipsoid, 2-seeded forms to c. 70 by 40 mm, without or with inconspicuous, oblique constriction, apex rounded. Seeds 1–2 per monocarp, 30–40 by 20–28 mm.
Peru (Loreto), fairly common in the region around Iquitos, not known elsewhere so far.
In forest on white sand. At elevations of 100–200 m. Flowering and fruiting: probably throughout most of the year.
Peru. Loreto: Mishana, 100–140 m, Ayala 1564 (AMAZ), Díaz S. et al. 404 (MO), Foster 4271 (F, MO, U), Gentry et al. 39313 (U), Vásquez et al. 5285 (MO, U), Vásquez & Jaramillo 9651 (MO, U); Puerto Almendras, 100–120 m, Díaz & Arévalo 81 (MO, U), Grández & Jaramillo 4983 (MO, U), Vásquez & Jaramillo 4619 (MO, U), 11031 (MO, U), Vásquez et al. 8067 (MO, U); Carretera de Peña Negra, at 2 km from Quista Cocha, 180 m, Rimachi Y. 4537 (MO, US), 7735 (MO, NA); Ninarumi (“Nina Rumy”), 123 m, Ruiz M. 808 (MO); Prov. Maynas, Allpahuayo, Vásquez et al. 17984 (MO).
http://species-id.net/wiki/Tetrameranthus_umbellatus
Figs 1A, B, G, 2F, G; Map 2Tunquí 62 (holotype U; isotype MO), Peru, Amazonas: Río Santiago, Huambisa, other side of La Poza 1 km, 180 m, 14 November 1979.
Tree, 8–25 m tall, 15–25 cm diam., young twigs and petioles densely to rather densely covered with stellate hairs <0.2 mm long, becoming glabrous. Leaves: petioles 5–25 mm long, 1.5–3 mm diam., lamina narrowly obovate or narrowly elliptic-obovate, 15–30 by 4–9 cm (index 2.4–4.7), chartaceous, greenish brown to brown above, pale greenish brown to brown below in sicco, sparsely covered with stellate hairs to glabrous above, rather densely to sparsely covered with stellate hairs on primary vein and sparsely covered to glabrous otherwise below, base acute to attenuate, decurrent along the petiole, apex acute to (abruptly) acuminate (acumen 5–10 mm long), primary vein flat to slightly impressed above, secondary veins 13–20 on either side of primary vein, slightly impressed to raised above, loop-forming, shortest distance between loops and margin 0.5–2 mm, tertiary veins flat to raised above, percurrent to more or less reticulate. Inflorescences up to 5-flowered, umbel-like; peduncles 5–25 mm long, c. 1.5 mm diam., densely to rather densely covered with stellate hairs, becoming glabrous in age, fruiting peduncles to c. 3 mm diam., bracts 4, oblong, 4–5 mm long, outer side densely covered with stellate hairs, falling before flowering, pedicels 25–70 mm long, ≥1 mm diam., densely to rather densely covered with stellate hairs, becoming glabrous in age, fruiting pedicels to c. 3 mm diam. Flowers green, maturing yellow in vivo; sepals broadly triangular-ovate to ovate, connate just at the base, to c. 4 by 3–5 mm, outer side densely covered with stellate hairs; outer petals ovate-elliptic, 16–20 by 9–12 mm, densely to rather densely covered with stellate hairs, the inner base with small glabrous area, inner petals elliptic, 13–17 by 6–7 mm, indument as in outer petals, the glabrous inner base slightly larger; stamens ≤1 mm long, connective shield cushion-shaped, flat. Monocarps 2–7, orange or red in vivo, brownish black in sicco, ellipsoid to oblongoid, 2-seeded forms to c. 45 by 20 mm and with oblique constriction, 1-seeded forms smaller and without constriction, mostly rounded at the apex. Seeds 1–2 per monocarp, to c. 25 by 18 mm.
Amazonian Peru (Amazonas, Huánuco, Loreto) and Brazil (Amazonas, Pará).
In non-inundated (terra firme) forest or rarely periodically inundated forest. At elevations of 100–600 m. Flowering and fruiting: probably throughout the year.
Brazil. Amazonas: Mun. Jutaí, Cid et al. 7269 (K, NY, US); km 1225 of Cuiabá-Santarém Highway (BR 163), Prance et al. 25602 (NY, U). Peru. Amazonas: Río Santiago, 2 km below Caterpiza, trail to Mitayar, E side of Quebrada Caterpiza, Huashikat 613 (MO, U); Quebrada Sasa, Monte Numi, 600 m, Kayap 2015 (MO, U). Huánuco: Distr. Llullapichis, Prov. Puerto Inca, Dantas, 280 m, Kröll Saldaña 712 (U); S of Pucallpa, next to the junction of the Río Pachitea and Río Llullapichis, 260 m, Morawetz & Wallnöfer 14-81085 (U); Prov. Pachitea, region of Pucallpa, Sira Mts., 26 km S of Puerto Inca, next to the junction of the Río Pachitea and Río Llullapichis, field station “Panguana”, 260 m, Morawetz & Wallnöfer 18-14188 (U, WU). Loreto: Prov. Maynas, Paucarillo Reserve, Río Amazonas, 110 m, Choo 347 (MO); Prov. Maynas, Distr. Las Amazonas, Explornapo Camp, near Sucusari, 100–140 m, Pipoly et al. 14565 (MO, U); Prov. Maynas, Llachapa, Río Napo, 130 m, Vásquez & Jaramillo 3732 (MO, U).
Peru: Washi yais (Tunquí 62), Wáshi yéis (Huashikat 613), Yaú (Kayap 2015).
The authors wish to thank three anonymous reviewers for critically reading of and commenting on the manuscript, and making a number of corrections. We very much thank Daniel Sabatier and Fanchon Prévost (CAY) for the discovery of Tetrameranthus guianensis in French Guiana, and for their efforts to collect additional fruiting and flowering material of it. We are indebted to the QCA Herbarium in Quito, for providing color slides of the rare Tetrameranthus globuliferus, and to Antonio Webber (Universidade do Amazonas, Manaus, Brazil) for photographs of Tetrameranthus duckei. We are very grateful to Roy Erkens (Utrecht University) for his comments on the phylogeny of the genus, and for preparing the distribution maps. We also want to thank Álvaro J. Pérez Castañeda who made some measurements on a flower preserved in alcohol in QCA, thus enabling to make the description of Tetrameranthus globuliferus more complete.