





(C) 2011 Leandro L. Giacomin. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new species of Solanum from Brazil is described. Solanum evolvuloides Giacomin & Stehmann, sp. nov. belongs to section Gonatotrichum, a small group assigned to the Brevantherum Clade of the genus Solanum. It resembles Solanum turneroides Chodat, sharing with it heterandry, and Solanum parcistrigosum Bitter, with which it shares a similar habit and pubescence. Despite these similarities, the species can be recognized by its ovate-elliptic to cordiform leaf shape and more membranaceous leaf texture than the other species in the section, and stem, inflorescence axes, and calyx vestiture mainly composed of glandular hairs. Solanum evolvuloides is known to occur only in southeastern of Bahia state, Brazil, and in a preliminary assessment of the IUCN criteria can be considered a threatened species.
ResumoUma nova espécie de Solanum é descrita para o Brasil. Solanum evolvuloides Giacomin & Stehmann, sp. nov. é componente da seção Gonatotrichum, um pequeno grupo associado ao Clado Brevantherum. A espécie é similar a Solanum turneroides, com a qual compartilha a heterandria, e Solanum parcistrigosum, que por sua vez apresenta um hábito e indumento foliar semelhante. Apesar da similaridade, a espécie pode ser facilmente reconhecida pela forma da folha ovada-elíptica a cordiforme e pela texura mais membranácea que as outras espécies da seção, além do indumento do caule, eixo da inflorescência e cálice, constituído em sua maioria por tricomas glandulares pedicelados. Solanum evolvuloides tem sua distribuição conhecida somente para o sudeste do estado da Bahia, Brasil, e é indicada como uma espécie ameaçada de extinção.
heterandry, Caatinga, IUCN conservation status
Solanum, with about 1500 species, is one of the largest genera of flowering plants, and the largest genus within Solanaceae (
Section Gonatotrichum was treated by
We revised material in the following herbaria (acronyms from Index Herbariorum, http://sweetgum.nybg.org/ih/): BHCB, CEPEC, CESJ, CTES, CORD, ESA, ESAL, FUEL, HAS, HB, HUEFS, IAC, IAN, ICN, INPA, LPB, MBM, MBML, PACA, R, RB, SP, SPF, SPSF, UEC, UPCB, VIC, and WU. Images of types deposited in BR, MO, NY, P, and SI, kindly provided by the curators or available on collections websites, were also examined. Plants obtained in the field were cultivated in the greenhouse and fresh flowers were fixed in alcohol to permit detailed descriptions and illustrations. We assessed the conservation status using IUCN Red List Categories and Criteria (
urn:lsid:ipni.org:names:77115891-1
http://species-id.net/wiki/Solanum_evolvuloides
Figs 1, 2Solano turneroidi Chodat et S. parcistrigoso Bitter similis sed ab utroque foliis membranaceis ovato-ellipticis vel cordiformibus et pilis glanduliferis in caule, axibus inflorescentiae, et calyce differt.
Brazil. Bahia: Mun. Jequié, Distrito de Cachoeirinhas, caatinga arbustiva em topo de morro, com lajeados graníticos, 299 m, 13°54'14.4"S, 40°01'46.8"W, 10 Jul 2009 (fr), L.L. Giacomin 974 (holotype: BHCB; isotypes, BM, MBM, NY, RB).
Herbs, slightly woody to woody at base, few- to many-branched, 20–40 cm tall. Stems moderately to densely pubescent with multicelled unbranched erect glandular hairs ca. 0.3–0.5 mm long, these mixed with less frequent slightly longer 1–3-celled unbranched eglandular hairs. Sympodial units difoliate, solitary or more commonly geminate, the smaller leaves up to half the size of the larger ones. Leaves simple, the blades 1–4 × 1–3 cm, ovate-elliptic to cordiform, chartaceous to membranaceous, sparsely to moderately pubescent on both sides with 1–2-celled unbranched erect eglandular hairs, these denser on the primary and secondary veins; venation camptodromous, with the primary and one pair of secondary veins emerging from the leaf base (sometimes just one, in the case of an asymmetric base), the primary and secondary veins barely visible to the naked eye, slightly prominent abaxially and less visible adaxially; base attenuate to cordate, slightly decurrent into petiole; margins entire, ciliate with hairs like those of the blade; apex acute to attenuate; petioles 0.5–2.2 cm long, with pubescence similar to that of the stems but with fewer eglandular hairs. Inflorescencessessile, lateral, extra-axillary or subopposite the leaves, unbranched, with 1–4 flowers, the axes with pubescence like that of the stems; peduncles absent; rachis very short; pedicels 6–10 mm in flower, 7–14 mm in fruit, almost contiguous, articulated at the base. Flowers 5-merous. Calyx 2–7 mm long, the tube 1–2 mm, the lobes 2–6 × 1–2.6 mm, ovate-elliptic, the apex acuminate, moderately pubescent abaxially with almost exclusively glandular unbranched multicellular erect hairs, densely pubescent adaxially with very small glandular hairs with 1-celled stalks; calyx accrescent in fruit, the lobes up to 8 mm long, equal to or exceeding the berry at maturity. Corolla 1–2.5 (-3) cm in diameter, rotate with abundant interpetalar tissue, membranaceous, white, the lobes 2–4 × 1–3 mm, triangular, acute at apex, with a few eglandular hairs abaxially, mainly on the central part of each lobe, glabrous adaxially. Stamens 4–9.5 mm long; filaments 1–2 mm long, with one much longer than the others, up to 5 mm long, glabrous; anthers 4–6 × 1.3–2 mm, connivent, yellow, the base cordate, with a small bulge dorsally, the apex emarginate, the pores directed introrsely and subapically, not opening into longitudinal slits. Ovary glabrous; style 7–9 mm long, longer than the smaller stamens, cylindrical, glabrous, curved near apex, closely appressed to the larger stamen; stigma capitate. Fruits 0.8–1.5 cm in diameter, globose berries, greenish white when immature, translucent at maturity, drying light-brown to blackish, glabrous, the mesocarp watery and held under pressure, dehiscing explosively at maturity, normally between two calyx lobes. Seeds 10–25 per fruit, 2.5–3.6 × 1.8–2.9 mm, flattened, reniform, with a small hollow where connected to the placenta, the margin flattened, forming a winged projection, the seed surface with raised projections and grooves parallel to margin, giving a netlike impression.
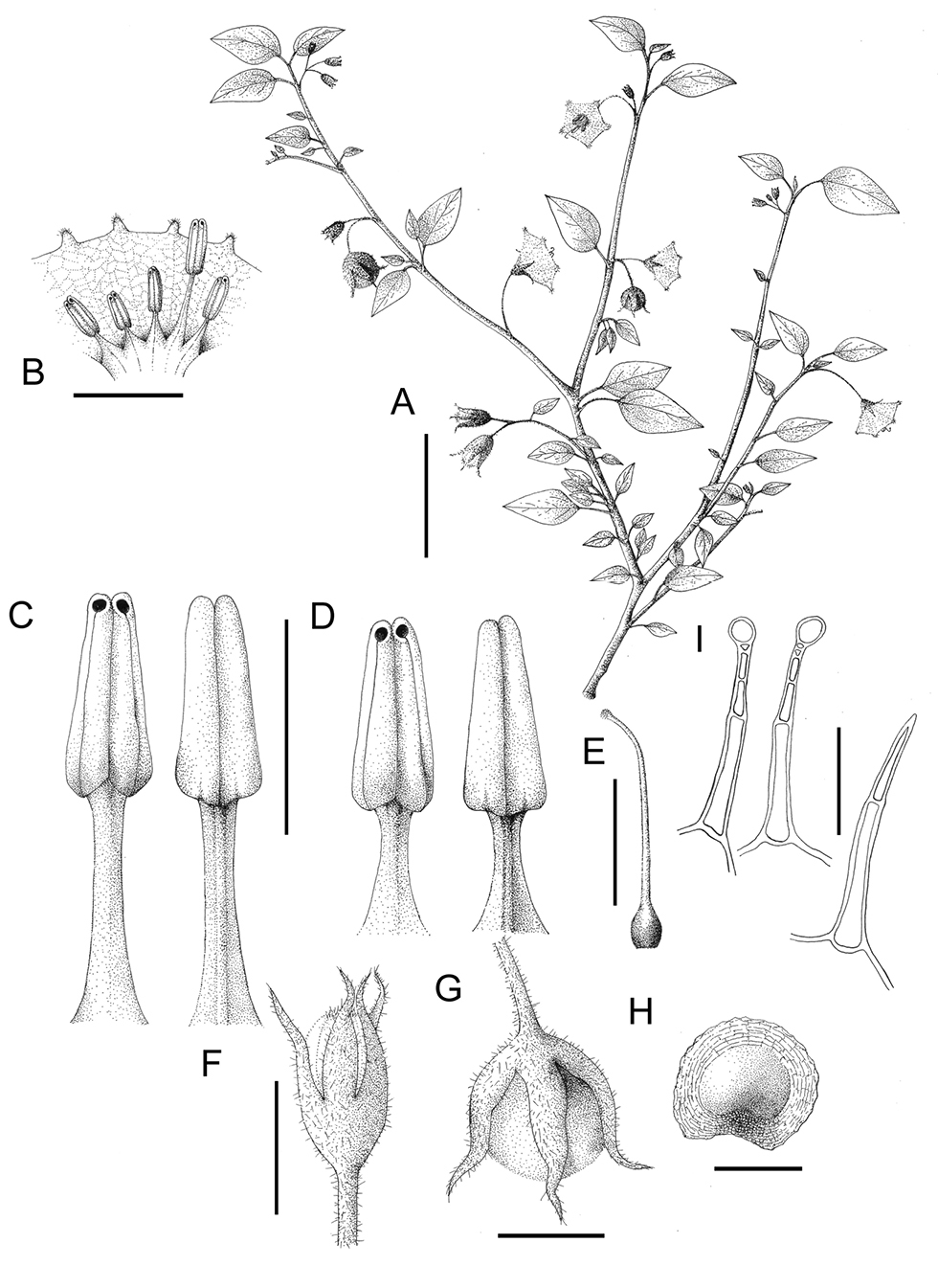
Solanum evolvuloides Giacomin & Stehmann. A Habit B Corolla cross section showing stamens C Larger stamen in ventral and dorsal view D Smaller stamens in ventral and dorsal view E Gynoecium F Young bud G Fruit H Seed I Detail of the hairs of stems, peduncles and calyces (glandular) and leaves (eglandular). All from Mattos-Silva s.n. (CEPEC-15698). Scale bars A= 3 cm; B, C, D, E, F and G = 5 mm; H = 2 mm ; I = 0.2 mm.

Solanum evolvuloides Giacomin & Stehmann. A Flower, opened during the morning in a cultivated seedling B Fruit, with a semi-transparent exocarp C Habit D Habitat in a shrubby caatinga formation. Scale bars A= 1.5 cm, B= 1 cm; C = 10 cm. All from Giacomin 974 (BHCB; type collection).
Distribution. This species is known only from the southeastern part of Bahia state (Fig. 3), Brazil, occurring in the transition zone between deciduous forests and xeric formations of shrubby Caatinga (as defined by
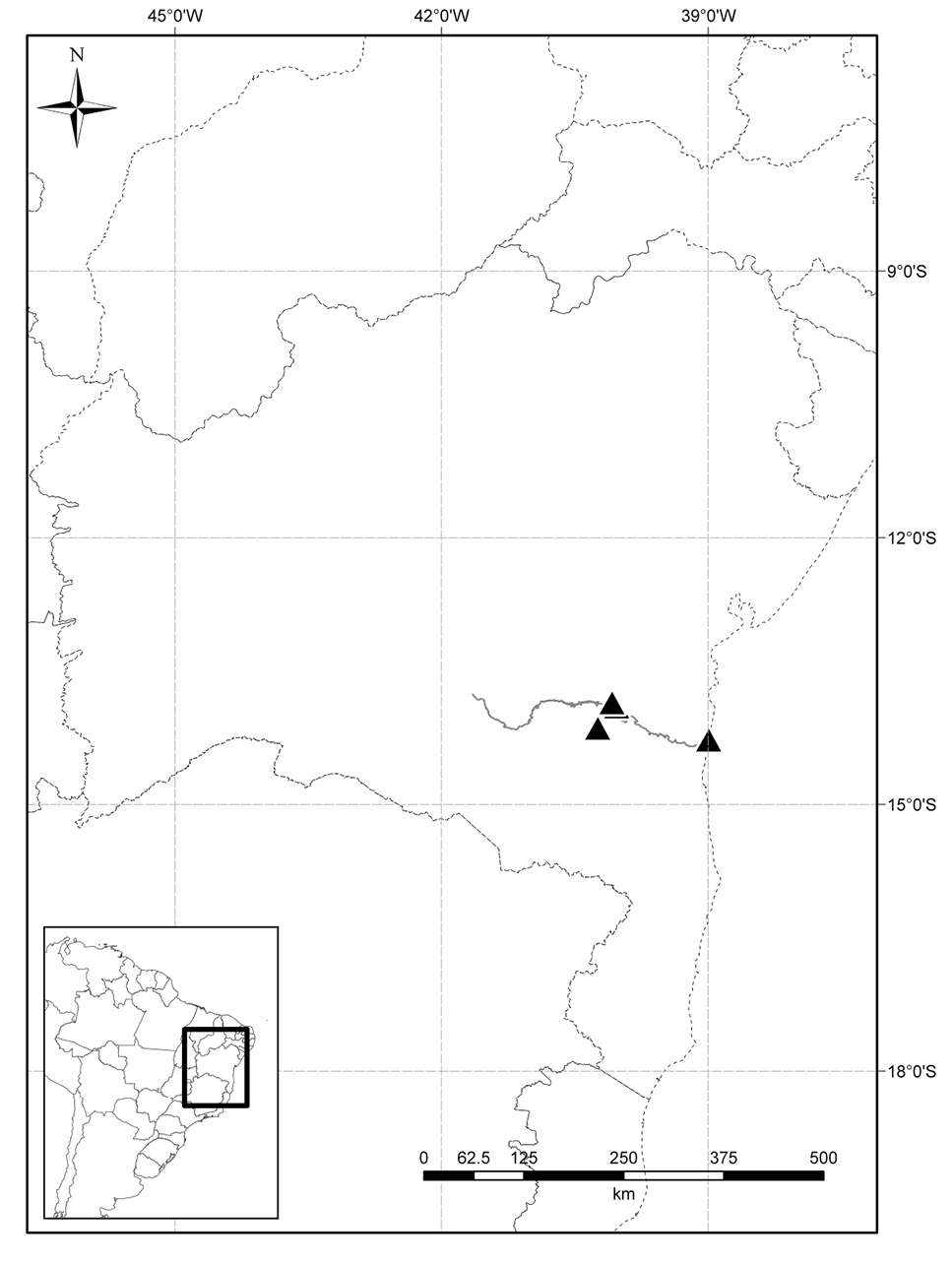
Known distribution of Solanum evolvuloides Giacomin & Stehmann (triangles) along Rio de Contas (in grey) in the state of Bahia, Brazil.
Ecology. Solanum evolvuloideswas recently recollected by the first author in the municipality of Jequié in a typical shrubby Caatinga formation, that is associated in this region with large granitic outcrops. The occurrence on the banks of the Rio de Contas near the city of Itacaré [Jardim, J.G. 1843 (CEPEC)] might be an occasional case of water dispersal by the river, which arises in a xeric environment near the center of the state in the Caatinga biome. Despite having been found in environments with marked seasonality, the species is apparently not annual, as evidenced by the woody stem bases.
Etymology. The name Solanum evolvuloides evokes the strong resemblance of this species to a sympatric species of the genus Evolvulus (Convolvulaceae).
Phenology. Flowering and fruiting material has been collected between February and August, with a flowering peak from February to May; fruiting specimens were collected from June to August. Under cultivation, flowers were observed to open only in the morning, closing during the evening. Observations of the same flower during consecutive days confirmed this pattern.
Preliminary conservation status. Endangered (EN) B1 a, b (i, ii, iii, iv). Solanum evolvuloides is known from only two localities, where the landscape has been strongly modified in the last decades due to the expansion of urban centers and extensive farming. The region has been focus of several surveys undertaken by the CEPEC group, in association with the New York Botanical Garden; despite this, only a few collections of this species have been made. Although one collection was made in a very disturbed area (Jardim 1843), the most recent collection is from a well-preserved forest fragment, and the species was not found in surrounding areas. There are no collections from within conservation units.
Specimens examined (Paratypes). BRAZIL. Bahia: Mun. Itacaré. Fazenda Monte Alegre, Ca. de 1 km a leste na rodovia para Itacaré. Margem do Rio de Contas, 10 August 1998 (fl, fr), J.G. Jardim 1843 (CEPEC); Mun. Jequié. Rodovia Ipiaú/Jequié, 12 May 1969 (fl, fr), J.A. Jesus 367 (CEPEC); Mun. Jequié. Km 7 da estrada Jequié/Ipiaú, Caatinga, 10 February 1983 (bs), A.M. Carvalho, 1591 (CEPEC); Mun. Jequié, Procedente do Distrito de Cachoeirinhas, caatinga arbustiva em topo de morro, com lajeados graníticos, 299 m, 13°54'14.4"S, 40°01'46.8"W, floresceu em cultivo no Jardim Botânico da Fundação Zoo-Botânica de Belo Horizonte, 23 September 2009 (fl), L.L. Giacomin 974B (BHCB, NY); Mun. Manoel Vitorino. Rod. Man. Vitorino / Caatingal. Km 4. região de Caatinga. 16 February 1979 (fl, fr), L.A. Mattos Silva s.n. (CEPEC [15698]).
Discussion. The species is similar to Solanum turneroides Chodatand they are the only species within the section presenting strong heterandry, with one stamen with a filament much longer than the other four. Sometimes Solanum parcistrigosum Bitterand Solanum hoffmannseggii Sendtner, species that also resemble Solanum evolvuloides, are weakly heterandrous but they both have smaller flowers (corolla with < 1.5 cm in diameter) and stamens, and can be easily distinguished from Solanum evolvuloides by the glandular indument observed in the calyx and stems of the latter species. This character can be also used to separate Solanum evolvuloides from Solanum turneroides. These two speciesare also not sympatric, in Brazil Solanum turneroides is found only in the states of Mato Grosso do Sul and São Paulo, whereas Solanum evolvuloides occurs only in Bahia. Solanum turneroides has an indument composed of unbranched eglandular hairs mainly with one-celled appressed hairs on the calyces, leaves and stems, associated with two-celled hairs that are typically geniculate (bent at a 90° angle) between the first and second cell. It is also a more robust shrub with chartaceous leaves, in contrast to the more membranaceous leaves of Solanum evolvuloides.
Within the section, glandular hairs are commonly found on the leaves of Solanum adscendens Sendtner but in this species, the hairs are much smaller (up to 0.1 mm, barely visible in dried material) than those of Solanum evolvuloides, have glands composed of more than one cell, and are associated with several erect eglandular hairs. The hairs of Solanum evolvuloides, perhaps the most distinctive characteristic of the species, are up to 0.5 mm long, are stalked, have unicellular glands, and are not commonly found on the leaves.
The heterandry found in Solanum evolvuloides is not a common feature within Solanum, and its evolution had been focus of recent studies, using morphological or molecular data (
We would like to thank L. Echternacht for assistance in the field and B. Radicchi for preparing the line drawings; T.E. Almeida for the GIS support; L. Bohs, S. Knapp, S. Stern and two anonymous reviewers for suggestions and critical review. This work was supported by FAPEMIG (APQ-01600-08) and CNPq (305589/2009). A Master’s scholarship was provided to LLG by CAPES.