





(C) 2011 Harold Robinson. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Nothovernonia gen. nov., is described as a new genus for the tropical African Vernonia purpurea Sch.Bip. ex Walp. and Vernonia amblyolepis Bak, having cymiform inflorescences, pedunculate heads with differentiated foliiform bracts at the base, apiculate involucral bracts with scarious lateral margins, spiculiferous corolla lobes, and strongly spinose, sublopohate tricolporate pollen with the colpi meeting at the poles. The new genus belongs to the subtribe Centrapalinae and a key to the known DNA-sequenced genera of the subtribe is provided. The new species names are Nothovernonia purpurea (Sch.Bip. ex Walp.) H.Rob. and V.A.Funk, comb. nov., and Nothovernonia amblyolepis (Baker) H.Rob. & V.A.Funk, comb. nov.
Compositae, Nothovernonia, new genus, tropical Africa, Centrapalinae, Erlangineae, phylogeny
Many genera have been named in the tribe Vernonieae over the last 200 years, see (
The species known as Vernonia purpurea is a coarse herb (Fig. 1) placed by
The goal of this study is to recognize the new genus and provide the proper nomenclature for a larger study on the subfamily Cichoroideae.
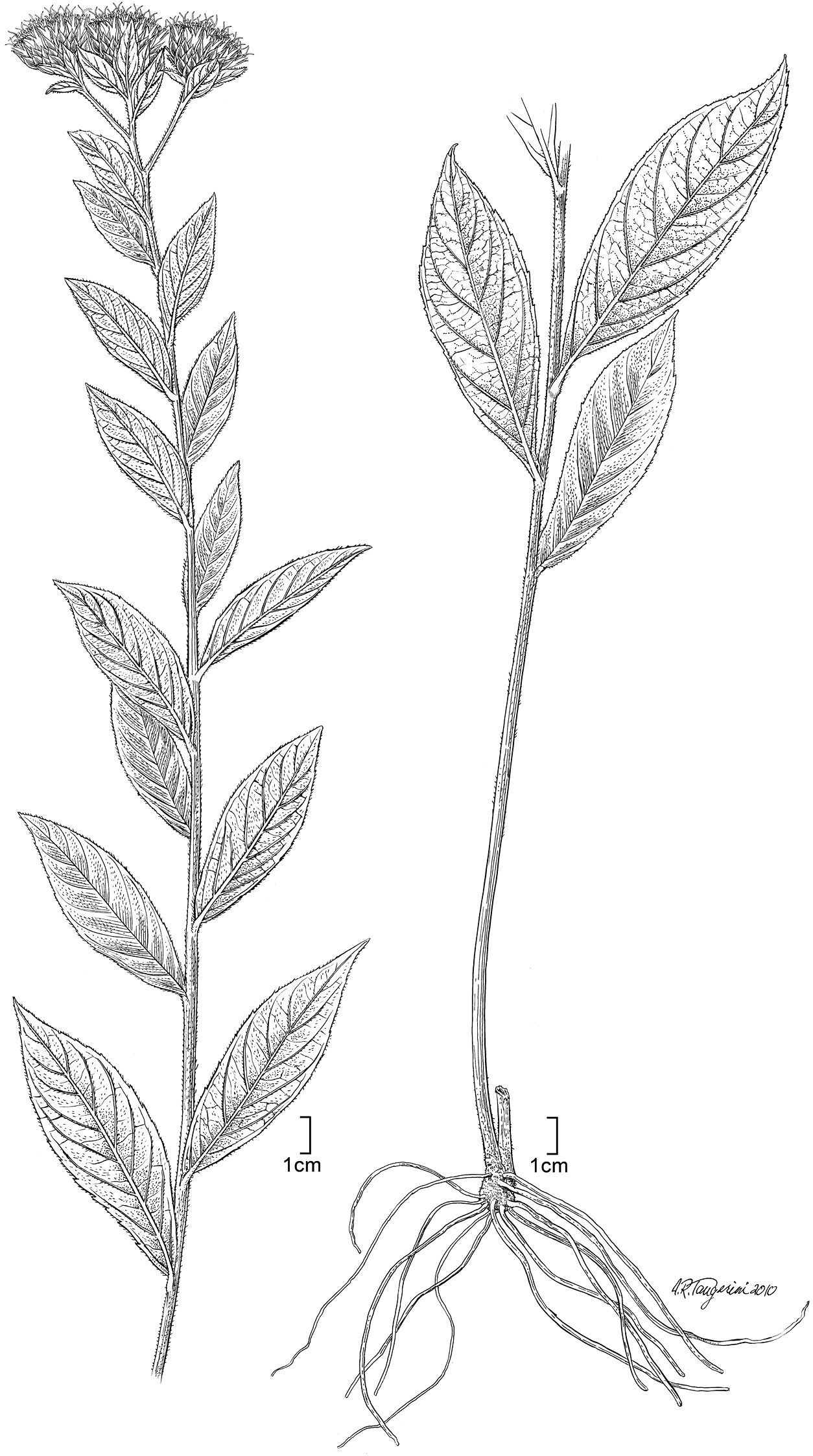
Nothovernonia purpurea (Sch.Bip. ex Walp.) H. Rob. and V.A. Funk: habit. [Illustration by Alice Tangerini (US)]
The morphology was studied using herbarium material,
most of which was from the U.S. National Herbarium in Washington, D.C.
Microscopic characters were examined via plant material mounted on
microscope slides in semi-permanent, water misable Hoyer’s Solution (
Scanned images of the syntypes of Vernonia amblyolepis Baker were sent by the Herbarium at the Royal Botanic Gardens, Kew.
The position on the molecular phylogeny was determined by analyzing DNA sequence data of ITS, ndhF, trnL-F and matK. The details of the molecular work will be published as part of a larger study of the subfamily Cichorioideae (
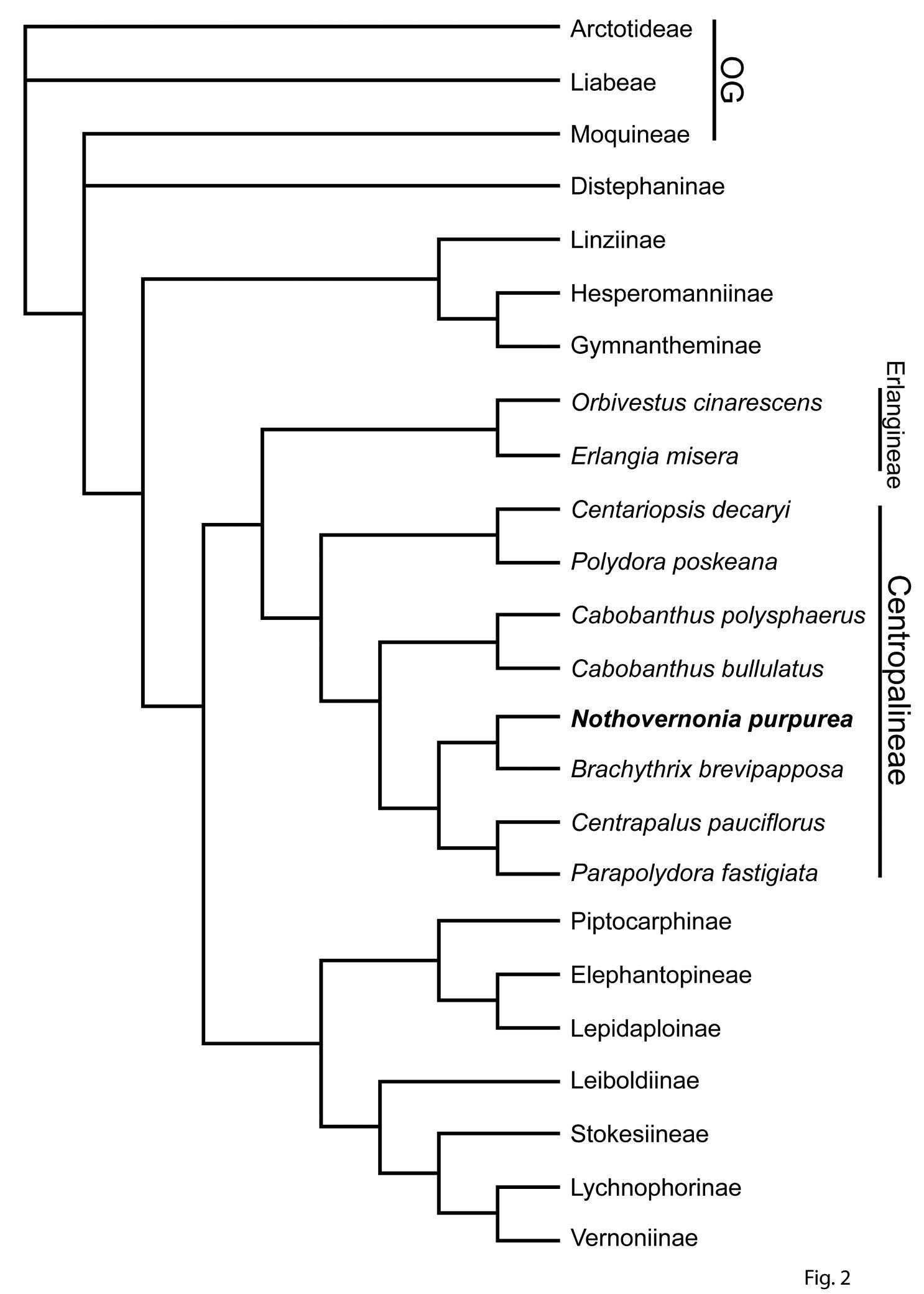
A phylogeny of the subtribes of the Vernonieae, with a detailed look at the subtribe Centropalineae. This tree is the strict consensus tree from a PAUP analysis that produced 28 trees. The data included were ITS, ndhF, trnL-F, and matK. The analysis included all known sequenced genera of the Centropalineae and is part of a larger analysis of the subfamily Cichorioideae.
The new genus, named here as Nothovernonia, can be distinguished from Centrapalus by the erect rather than decumbent bases of the stems, the apiculate laterally scarious rather than long herbaceous non-scarious tips on the involucral bracts, and the purple or lilac rather than blue-purple corollas (Fig. 3, B-D). The corolla color may be most useful in the field, with Centrapalus usually being bluish and Nothvernonia apparently never being blue. The stout spreading sweeping hairs of the style branches are different from the slender usually more appressed hairs in Centrapalus. The achenes are similar in having narrow though small raphids, many idioblasts on the surface, setulae with pairs of cells usually separated to a third or more of their length, but differ in the cells of the setulae being fused in the basal one to two thirds and the pappus being white to rufous. In Centrapalus the cells of the setulae are separate and essentially solitary to near the base, and the pappus is sordid grayish.

Images of Nothovernonia purpurea (Sch.Bip. ex Walp.) H. Rob. and V.A. Funk A Habitat B Head before flowering with outer bracts tightly appressed C Fully flowering head showing well developed bracts D Head with only a few flowers but showing the well developed bracts. [Photographs by A. Thiombiano, M. Schmidt, and K. Schumann]
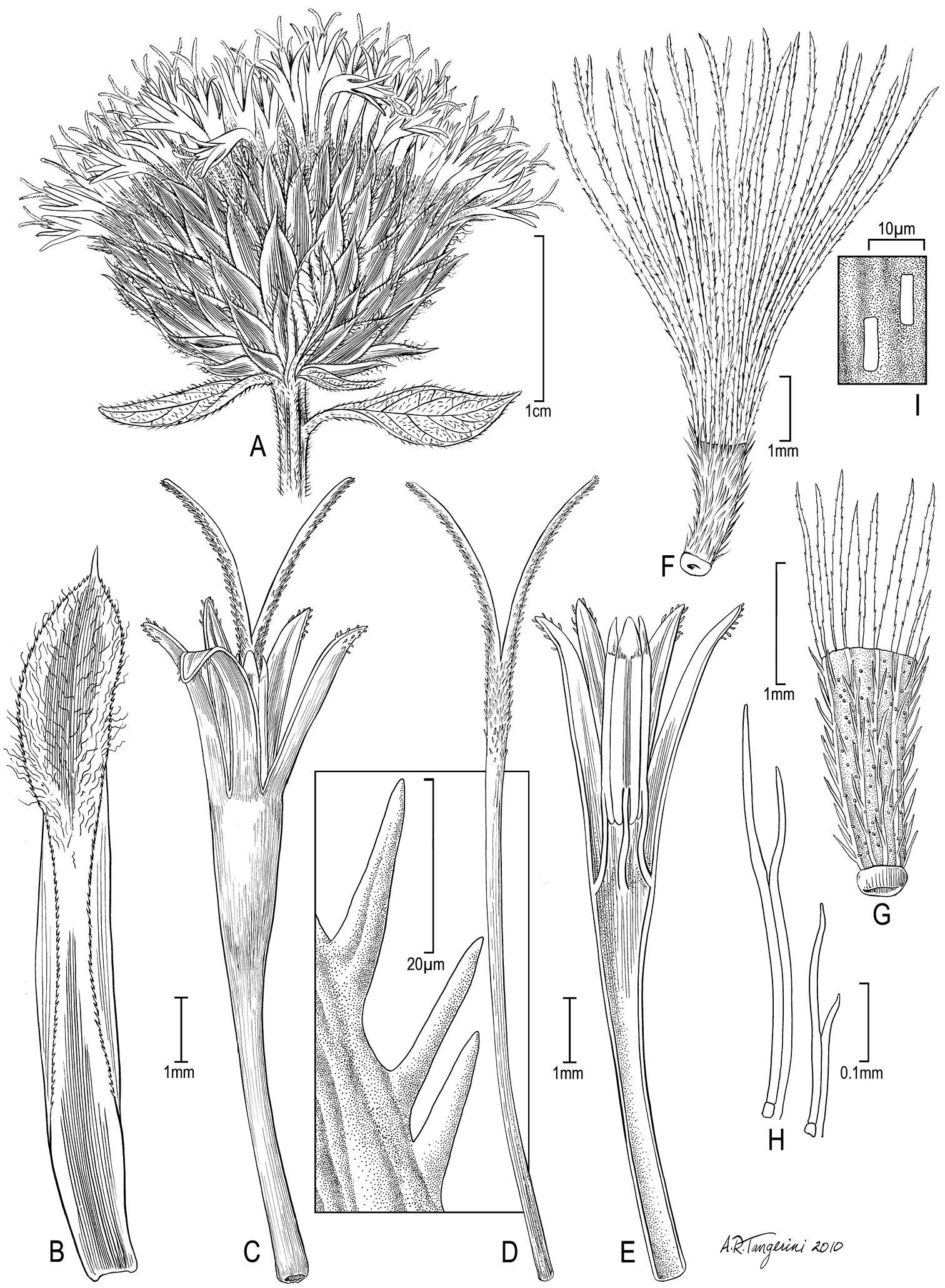
Nothovernonia purpurea (Sch.Bip. ex Walp.) H. Rob. and V.A. Funk A head B involucral bract C floret with style branches D style branch with detail of sweeping hairs in square E longitudinal section of floret showing anthers F achene with pappus G achene H detail of achene setulae I examples of raphids from achene wall. [Illustration by Alice Tangerini (US)]
Robinson and Skvarla published images of the pollen of Nothovernonia (
The two species treated here have their corolla color and stem bases more like those of Vernonella, but the colpi of the pollen are not truncated and thus failing to reach the poles, the involucral bracts are not scarious across the tips, the corolla lobes are spiculiferous with stiff hairs outside distally, and the achenes have setulae with cell pairs separated nearly half way but not to the base. In addition, the two species differ from both Centrapalus and Vernonella by the small to large differentiated foliiform bracteoles at the base of the head. This contrasts with the loose but undiffentiated narrow involucral bracts at the base of the head in Centrapalus and the small and broad but otherwise undifferentated bracts at the base of the head in Vernonella.
On the basis of structural features, the genus is clearly distinct, but the position nearer Centrapalus of the subtribe Centrapalinae or closer to Vernonella of the Linziinae, has been resolved with certainty only with DNA sequencing. Sequence data are available for Centrapalus, Parapolydora and Nothovernonia purpurea (see below). Members of Vernonella
have not been sequenced, but the subtribal placement is derived from
the report of elemanolide sesquiterpene lactones from one of the
species, Vernonia praemorsa (Muschl.) H.Rob. & Skvarla (
Figure 2
is the result of the analysis of DNA sequence data and shows all of the
subtribes in the Vernonieae and all the known genera within the Centrapalinae. Within this scheme, Vernonia purpurea belongs in the broad group first placed by
Genera known to be in the subtribe Centrapalinae based on DNA sequence data are shown in Figure 2.
Key to the Genera known to be in the Centrapalinae(for a key to the subtribes of Vernonieae see
| 1 | Receptacles with paleae | Centauriopsis |
| – | Receptacles without paleae | 2 |
| 2 | Inner pappus reduced of 3–15 short easily caduous bristles | Brachythrix |
| – | Inner pappus of 30 or more well-developed rather persistent long capillary bristles | 3 |
| 3 | Inflorescence spiciform, with clusters of heads in axils of leaves | Cabobanthus |
| – | Inflorescence a spreading panicle | 4 |
| 4 | Heads 0.5 cm or less wide, with 12–30 florets; setulae of achene with paired cells united essentially to tip | 5 |
| – | Heads 0.5 cm or more wide, with 30 florets or more; setulae of achenes with paired cells distinctly separated at tips or from base | 6 |
| 5 | Plants mostly annuals; stems with simple or L-shaped hairs pappus bristles usually tawny, yellowish or green, rarely white; pollen triporate or with short colpi | Polydora |
| – | Perennials; stems, leaves, involucres and corolla lobes with large T-shaped hairs; heads with 12–20 florets; pappus white; pollen sublophate, tricolporate | Hilliardiella |
| 6 | Setulae of achenes with pairs of cells fused to a third or more of their length; heads with small or large foliose bracts at base; involucral bracts acuminate at tips | Nothovernonia |
| – | Setulae of achenes with cells separated or solitary for most of their length; heads without differentiated foliose bracts at base; involucral bracts mostly with narrowly attenuate tips | 7 |
| 7 | Base of stem decumbent; pappus bristles grayish or tawny; setulae of achenes with cells elongate but separated to near base | Centrapalus |
| – | Base of stem erect; pappus bristles whitish; setulae of achenes with only one of cells elongate and solitary from near base | Parapolydora |
Ad Vernonella in habitis herbaceis et floribus purpureis simila sed capitulis base bracteoliferis in lobis corollarum distaliter spiculiferis et in grana pollinis sublophatis totaliter tricolporatis differt. A Centrapalus in basis erectis et bracteis involcri apiculatis lateraliter scariosis differt.
Vernonia purpurea Sch.Bip. ex Walp.
Coarse branching herbs to 0.7 m tall, stems erect from base, distinctly ribbed, pilose with spreading simple hairs. Leaves alternate with petioles 0.2–2.0 mm long; leaf blades oblong to lanceolate, sparsely pilose above with prominent persistent bases on the hairs, lower surface coarsely and densely pilose on major veins, surface with many glandular dots, secondary veins pinnate, ca. 6 pairs. Inflorescences terminal and from axils of reduced upper leaves, distinctly cymiform with distinct short to long peduncles; heads broadly campanulate, with minute to large foliose bracts at base; involucral bracts in ca. 5 series, strongly gradate, appressed, ovate to narrowly oblong, with dark median stripe, apex apiculate with distinct dark or rarely pale awn, with pale scarious lateral margins, outer surface with numerous arachnoid hairs from median band, spreading tranversely as the head expands; receptacles flat, without pales or hairs. Florets ca. 30–65 per head; corollas purple, with lobes and upper throat exceeding the pappus at anthesis, glanduliferous on throat, lobes spiculiferous with stout straight hairs distally on outer surface; anther thecae with small sterile border at base, endothecial cells oblong with sinuous lateral walls, apical appendage triangular, firm. Style base with annuliform sclerified node, distally with stout spreading sweeping hairs covering backs of style branches and upper 1 mm of style shaft. Achenes prismatic, 8–10-ribbed, with numerous idioblasts on surface, with small narrow raphids, setulae with pairs of cells fused together beyond basal 1/3; pappus with inner series of many crowded capillary bristles, white or rufous, less than 2/3 as long as corollas; outer pappus series of shorter, crowded lanceolate scales.
Pollen grains ca. 40 mm in diam., tricolporate with colpi reaching poles, spinose, surface sublophate with perforated tectum continuous between colpi, bacculae single under each spine.
Chromosome number n = 9 (
The new generic name, Nothovernonia, means “false Vernonia”.
urn:lsid:ipni.org:names:77111572-1
http://species-id.net/wiki/Nothovernonia_purpurea
The species is known from the Sudan, Ethiopia, Kenya, Tanzania, and Uganda, south to Angola and Malawi and west to southern Senegal.
Congo. Bafuka (ueli), 1929, Steyaert77(US). French Equatorial Africa. Dans la Haute-Kotto (Oubangu-Chari-A.E.F.), 100 km NW Labuya, 1921–1923, herb. G. le Testu 4127(BM, US). Malawi. s.l., 1891, Buchanan 143 (K, US). Machinga District: Liwonde Forest Reserve, in Brachystegia woodland on steep hillside, 15°07'S, 35°23'E, 3000 ft., 11 Apr 1984, Christenson and Solubeni 1454(US). Ntcheu District: Golomati Road, 4 km E of road to Dedza, 14°50'S, 35°25'E, 20 Apr 1984, Christenson, Patel and Lipende 1477 (US). Lilongwe District: Dzalanyana Forest Reserve, about 8 km from entrance gate, 14°15'S, 33°25'E, 3000 ft, 2 May 1984, Christenson and Lipende 1493(US). Senegal. Tambacounda, Dindéfello, à proximité du campement touristique, dans le vallée de la cascade, 12°22'N, 12°19'W, 200 m, 22 Sep 1994, Sambou et Madsen, Goudiaby, Traoré and Laegaard 319 (AAU, DAKAR, US). Uganda. s.l., s.d., Dummer 84(US, isotype of Vernonia dummeri). Upper Volta. Ifan, Solenso, 16 Sep 1974, Bognounou-Quattara 2(US). Zambia. Kitwe, 15 May 1967, DZF? F10, 062(NDO, US).
Figure 3 illustrates the habit of Nothovernonia purpurea, note the large bracteoles at the base of the capitula. Figure 4 has the details of the capitula, note the stout hairs of the style branches (Fig. 4D) and the setulae of the achenes that have pairs of cells fused together beyond the basal 1/3 (Fig. 4H).
Nothovernonia purpurea has large, obvious foliiform bracteoles at the bases of the capitula, bracteoles that can often cover the involucre completely. In contrast, Nothovernonia amblyolepis has minute foliose bracts that are easily overlooked.
urn:lsid:ipni.org:names:77111573-1
http://species-id.net/wiki/Nothovernonia_amblyolepis
The species is known from Angola, Malawi, Rwanda and Tanzania.
Malawi. Zomba District: Zomba Plateau, top of hillside above KuChawe Inn, on grassy slope, 15°20'S, 35°18'E, 5500 ft., 14 Apr 1984, Christenson 1461 (US). Zomba District: Zomba Plateau, on upper road from KuChawe Inn downward to Zomba, near horse paddock, 15°20'S, 35°18'E, 5000 ft., 25 Apr 1984, Christenson and Lipende 1488(US). Uganda. Buhweju County: District West Ankole, Nyarwambu River, 0°22'S, 30°28'E, 1550 m, 23 Aug 1982, Rwaburindora 919(MO, US).
| Inflorescence copious, with a number of branches arising from the uppermost leaf-axils; ultimate peduncles short, so that the capitula appear to be in clusters | 62. Vernonia amblyolepis | |
| Inflorescence strictly terminal, few-headed, lax, the ultimate peduncles longer, so that the capitula do not appear to be in clusters | 61. Vernonia kandtii |
The specimens from Malawi and Uganda that were examined for this study show variation in the structure of the inflorescence and therefore, do not support a separate species status for Vernonia kandtii.
We thank Alice Tangerini (US) for the beautiful illustration and Stan Yankowsky who did microphotography to aid in the illustration. The Royal Botanic Gardens, Kew kindly sent us scans of the syntypes. Funk thanks Raymund Chan for allowing her to use a small part of the preliminary results of their Cichorioideae study. For the photographs we thank Prof. Adjima Thiombiano, Ouagadougou University, Dr. Marco Schmidt, Senckenberg Research Institute, and Katharina Schumann, Goethe-University Frankfurt. This and other west African plants can be found on their website www.westafricanplants.senckenberg.de.