





(C) 2010 Sandra Knapp. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Three new species of “non-spiny" Solanum are described from Peru and Ecuador, and a revised description for Solanum verecundum M. Nee is presented. Solanum kulliwaita S. Knapp, sp. nov. (Dulcamaroid clade) is endemic to the Department of Cuzco in southern Peru, and is most similar to the recently described Solanum sanchez-vegae S. Knapp of northern Peru. Solanum dillonii S. Knapp, sp. nov. (Brevantherum clade) is found in southern Ecuador and northern Peru in the Amotape-Huancabamba phytogeographic zone, and is morphologically similar to the widespread Solanum riparium Ruiz & Pav. Solanum oxapampense S. Knapp, sp. nov., (also of the Brevantherum clade) is endemic to the Oxapampa region (Department of Pasco) of central Peru, and is similar to and segregated from Solanum verecundum M. Nee of Peru and Ecuador. Complete descriptions, distributions and preliminary conservation assessments of all new species are given.
Amotape-Huancabamba zone, Andes, Ecuador, endemism, conservation, nightshades, Peru, Solanum
Solanum L., with ca. 1500 species, is the largest genus in the Solanaceae and one of the ten most species-rich genera of flowering plants (
Recent intensive collecting in Peru, coupled with
targeted collecting by members of the PBI project team and work in many
herbaria has uncovered several new species from Peru and adjacent
Ecuador and necessitated the revision of the circumscription of Solanum verecundum
M. Nee from which one of these new taxa is segregated. Two of these are
endemic to Peru and one to the recently defined highly diverse
Amotape-Huancabamba phytogeographic zone (
Members of the Dulcamaroid clade (sensu
Species nova Solano sanchez-vegae mihi similis, sed foliis ad apicem acuminatis, inflorescentibus et floribus glandularibus, trichomatibus uniseriatis simplicibus differt.
Peru: Cusco: Prov. La Convención, Dist. Ocobamba, Mesa Pelada, 12°54.13S, 72°37.06W, 2613 m, 23 March 2004, L. Valenzuela, E. Suclli & G. Calatayud 3163 (holotype: USM!; isotypes: AMAZ, CUZ, MO!, MOL, NY! [NY00824906]).
Woody vine or scandent shrub, height unknown, the branches arching. Stems sparsely pubescent with simple uniseriate multicellular trichomes 0.5–1 mm long, glabrescent, slightly winged from the decurrent leaf bases; new growth pubescent with simple or occasionally branched uniseriate trichomes 0.5–1 mm. Bark of older stems dark reddish brown, shiny. Sympodial units plurifoliate. Leaves simple, (2-)3.5–8.5 cm long, 1–3 cm wide, narrowly elliptic to lanceolate, slightly fleshy, the upper surfaces sparsely pubescent with simple or occasionally furcate or branched trichomes on the lamina, more densely pubescent on the midvein, the lower surfaces glabrous or with a few scattered simple uniseriate trichomes along the midvein; primary veins 7–9 pairs, often drying blackish brown; base acute to attenuate; margins entire, sometimes revolute, densely pubescent in the basal quarter to third with simple trichomes extending from the petiole; apex acute; petioles 0.7–2 cm long, densely pubescent along the adaxial groove with golden simple or occasionally furcate uniseriate trichomes, not apparently twining. Inflorescences terminal or appearing lateral, 9–11 cm long, 3–5 times branched, with 10–20 flowers, densely pubescent with simple uniseriate trichomes mostly 0.3–0.5 mm long, some longer and to 1 mm, purple in live plants and retaining pigmentation in dried material, the cells of the trichomes small and weak-walled, usually collapsing and tangled, the lateral cell walls dark-pigmented, the terminal cells spheroidal and apparently glandular; peduncle 1.5–3.5 cm long; pedicels 1–1.2 cm long, ca. 0.5 mm in diameter at the base, ca. 1 mm in diameter at the apex, slender, erect to nodding, densely pubescent like the inflorescence axes, articulated at the base and inserted into a short sleeve or above the base and leaving a peg ca. 2 mm long; pedicel scars irregularly spaced 0.5–5 mm apart, usually grouped. Buds ellipsoid, the corolla strongly exserted from the calyx tube before anthesis. Flowers all perfect, 5-merous. Calyx tube 2–2.5 mm long, cup-shaped, narrowing gradually to the pedicel, the lobes 2.5–3.5 mm long, the lower portion broadly deltate, the distal part an apiculate tip to 2 mm long, densely pubescent with simple uniseriate trichomes like those of the inflorescence axes abaxially, these apparently glandular, the adaxial surface glabrous. Corolla 2.3–2.5 cm in diameter, purple, stellate, lobed 2/3 to ¾ of the way to the base, the lobes 9–12 mm long, 4–5 mm wide, spreading, the tips and margins densely pubescent on the abaxial surface with weak, collapsing simple uniseriate trichomes like those of the inflorescence, but smaller and not apparently glandular. Filament tube minute, the free portion of the filaments 1–2 mm long, glabrous; anthers 3.5–4.5 mm long, 1–1.5 mm wide, ellipsoidal, loosely connivent, yellow, poricidal at the tips, the pores lengthening to slits with age. Ovary glabrous; style 7–8 mm long, glabrous; stigma capitate, the surface minutely papillose. Fruit a globose berry, ca. 1 cm in diameter (immature?), black when ripe, the pericarp thin, not shiny, glabrous; fruiting pedicels 1.5–1.7 cm long, ca. 1.5 mm in diameter at the base, woody, more or less nodding. Seeds not known.

Figure 1. Isotype specimen of Solanum kulliwaita S. Knapp. (Valenzuela et al. 3163, NY [NY00824906]). Specimen image reproduced with the permission of The C. V. Starr Virtual Herbarium of The New York Botanical Garden (http://sciweb.nybg.org/science2/VirtualHerbarium.asp).
Endemic to the valley of the Río Urubamba in the Department of Cusco in southern Peru (Fig. 2).
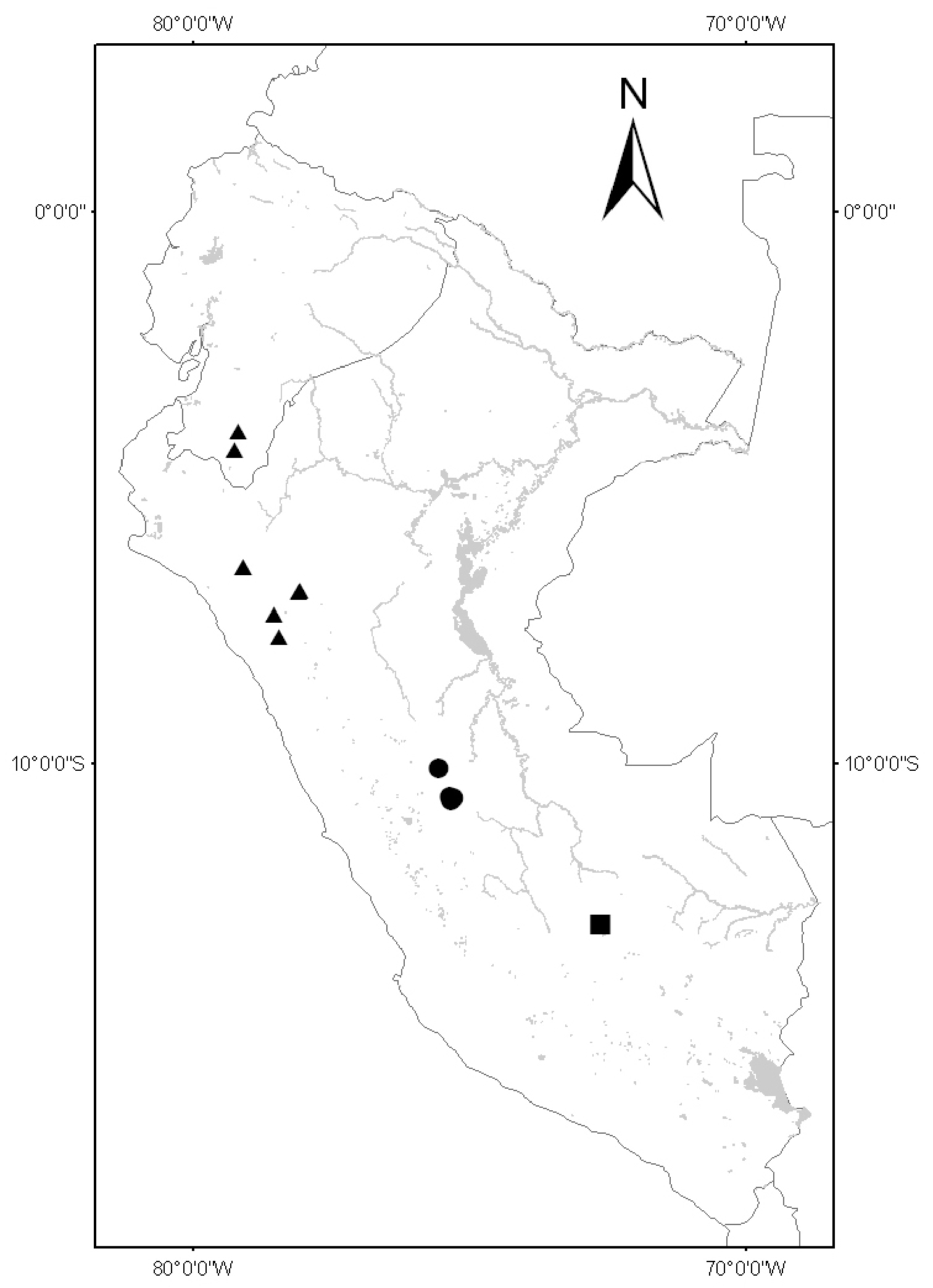
Figure 2. Distribution of Solanum kulliwaita (circles), Solanum dillonii (triangles) and Solanum oxapampense (squares).
Both collections are from montane cloud forests of the eastern slopes downriver from Machu Picchu in a locality broadly known as Mesa Pelada, from 2400–2600 m.
Named for the flower and trichome colour – kulli = purple; waita = flower in Quechua.
Known only from two collections in a very narrow geographical area outside any protected area, Solanum kulliwaita
is assessed as Data Deficient for EOO and AOO (due to there not being 3
collection points with which to calculate the polygon: Moat, 2007);
using the cell size of 2 km2, however, is assessed as Critically
Endangered (
Peru: Cusco: Prov. La Convención, Dist. Maranura, Mesapelada, 12°54.33S, 72°37.06W, 2450 m, 19 April 2004, W. Galiano, E. Suclli, P. Núñez, A. Rodriguez & V. Chama 6137 (CUZ, MO, NY! [NY00824933], USM).
Solanum kulliwaita is most similar morphologically to Solanum sanchez-vegae S. Knapp from northern Peru; both species have large, purple flowers and simple leaves. It can be distinguished from the latter species by its leaves that are glabrous beneath and with a dense covering of uniseriate trichomes on the adaxial surface of the petiole, the ciliate lower leaf margins and the distinctive uniseriate glandular trichomes of the inflorescence. Solanum sanchez-vegae has larger, fleshier leaves with loose dendritic trichomes and the trichomes of the inflorescence are both non-glandular and dendritic. The inflorescence trichomes of Solanum kulliwaita are unusual in members of the Dulcamaroid clade in having three globular cells at the apex and in drying purple (and being purple in live plants, fide Valenzuela et al. 3163). Trichomes on the rest of the plant (i.e., leaves and stems) are not glandular.
Brevantherum clade
Members of the Brevantherum clade (sensu
Species nova Solano ripario Ruiz & Pav. similis, sed trichomatibus multangulatis vel echinatis longistipitatis, foliis ad basibus acutis, floribus violaceis, differt.
Peru: Cajamarca: Prov. Celendin, on road from Celendin to Balsas, east of pass on descent to Balsas, 2002 m, 6°52.13S, 78°30.91W, 12 December 2007, S. Stern, E. Tepe, S. Leiva & M. Zapata 119 (holotype: USM!; isotypes: BM! [BM001016881], HAO†, NY! [NY00986687], UT!).
Shrub or small tree, 4–8 m tall, branching in the upper part of the stems. Stems densely pubescent with multangulate to echinoid trichomes on multiseriate stalks 0.5–1 mm long, the rays > 12, 0.2–0.3 mm long, glabrescent; new growth densely pubescent with multangulate to echinoid trichomes on multiseriate stalks 0.5–1 mm long like those of the stems, greyish white. Bark of older stems reddish brown. Sympodial units plurifoliate, the branching dichasial. Leaves simple, 12–30 cm long, 4.5–16 cm wide, elliptic to broadly elliptic, discolorous, the upper surfaces evenly and moderately pubescent with 1–3-rayed sessile stellate trichomes, the rays 0.3–1 mm long, the trichome bases bulbous, the lamina clearly visible, the lower surfaces densely pubescent with of multangulate to echinoid trichomes on multiseriate stalks 0.5–1.5 mm long, the rays 10–16, to 1 mm long, these mixed with porrect-stellate trichomes with 8–10 rays on multiseriate stalks to 1 mm long, and sessile echinoid trichomes with weak rays to 0.3 mm long, the lamina not visible; primary veins 9–11 pairs, the veins drying yellowish green above, not visible beneath; base acute; margins entire, plane; apex acute; petioles 1.5–3 cm long, densely pubescent with multangulate to echinoid trichomes like those of the stems and leaf undersurfaces. Inflorescences terminal, 15–20 cm long, many times branched, with 100+ flowers, densely pubescent with multangulate trichomes of many sizes, the largest to 0.l8 mm in diameter, on multiseriate stalks to 1.5 mm, some smaller and sessile; peduncle 7–10 cm long; pedicels 7–9 mm long, ca. 2 mm in diameter at the base, 2.5–3 mm in diameter at the apex, stout, nodding at anthesis, densely pubescent like the inflorescence axes, articulated at the base; pedicel scars closely spaced ca. 1 mm apart. Buds globose, the corolla scarcely exserted from the calyx tube before anthesis. Flowers all perfect, 5-merous. Calyx tube 3–3.5 mm long, cup-shaped, narrowing gradually to the pedicel, the lobes 2.5–3 mm long, deltate, densely pubescent abaxially with multangulate to echinoid trichomes like those of the inflorescence rhachis, these more sessile distally, the adaxial surface sparsely pubescent with sessile echinoid trichomes. Corolla 1.3–1.5 cm in diameter, purple, stellate, lobed ca. ¾ of the way to the base, the lobes 6–7 mm long, 3.5–4.5 mm wide, reflexed or spreading at anthesis, the tips and margins densely pubescent on the abaxial surface with sessile or short-stalked multangulate to echinoid trichomes with >10 rays like those of the inflorescence, the adaxial surface glabrous or with a few echinoid trichomes near the apex on the midvein. Filament tube minute, the free portion of the filaments 2.5–3 mm long, glabrous; anthers 3–3.5 mm long, ca. 1 mm wide, ellipsoidal, loosely connivent, yellow, poricidal at the tips, the pores lengthening to slits with age. Ovary densely pubescent with multangulate trichomes; style 9–9.5 mm long, sparsely pubescent along its entire length with multangulate trichomes with 4–8 rays; stigma clavate, the surface minutely papillose. Fruit a globose berry, 1–1.5 cm in diameter, dark green when ripe, the pericarp thin, not shiny, unevenly pubescent with sessile or short-stalked multangulate trichomes with rays of many varying lengths, the longest rays ca. 1 mm long; fruiting pedicels 1.2–1.5 cm long, ca. 3 mm in diameter at the base, woody, more or less erect. Seeds >200 per berry, 1.5–2 mm long, 1–1.5 mm wide, flattened-reniform, reddish or golden brown, the surfaces minutely pitted.

Figure 3. Isotype specimen of Solanum dillonii S. Knapp. (Stern et al. 119, NY [NY00986687]). Specimen image reproduced with the permission of The C. V. Starr Virtual Herbarium of The New York Botanical Garden (http://sciweb.nybg.org/science2/VirtualHerbarium.asp).
Southern Ecuador (Prov. Loja) and northern Peru (Dept. Cajamarca), in the Amotape-Huancabamba phytogeographic zone (see
Tropical moist forest along the western slopes of the Andes and the valley of the Río Marañon, from 1500–2200 m. Often found along roads and small streams in secondary situations.
Named in honor of Michael O. Dillon, recently retired from the Field Museum of Natural History in Chicago, who has devoted much time and energy to the understanding of the flora of northern Peru and whose efforts collecting in the Monteseco region helped define the extent of this species' distribution.
Solanum dillonii
is relatively widely distributed in the Amotape-Huancabamba region,
and has an EOO of 22, 492 km2, giving a status of possible Near
Threatened, and an AOO (with cell size of 0.04 km 2) of 10, 541 km2,
giving a status of Least Concern (
Ecuador: Loja: Vilcabamba-Yangana road km 1–3, 1500–1600 m, 4°18'S, 79°14'W, 29 October 1984, P.M. Jørgensen 56267 (AAU, BM [BM001012348], QCA); ca. 7 km E of Catamayo on the road to Loja, ca. 1700 m, 3°28'S, 79°10'W, 6 February 1984, S. Knapp & J. Mallet 6258 (BH, QCA, NY [NY00829072], US). Peru: Cajamarca: Prov. Santa Cruz, Dist. Catache, upper Río Zaña valley, ca. 5 km above Monte Seco on path to Chorro Blanco, 1500–2000 m, [6°52'S, 79°05' W], 16–18 March 1986, M.O. Dillon, A. Sagástegui A., D. Dillon, P. Alcorn, J. Santisteban, S. Leiva, C. Téllez & M. Guzmán 4379 (BM [BM000849416], F [F-1993682], NY [NY00829133]); Prov. Santa Cruz, ca. 3 km por aire ENE Monteseco, 1800 m, [6°52'S, 79°05'W], 31 May 1987, J. Santisteban C. & J. Guevara B. 125 (BM, NY [NY00829131]); Prov. Celendin, Marañon River valley, Chachapoyas-Cajamarca road, 1900–2100 m, [6°51'S, 78°04'W], 28 May 1984, D.N. Smith & J. Cabanillas 7257 (MO [MO-5294201], NY [NY00829129]); Prov. Cajamarca, Dist. San Juan, 2 km before town of San Juan, on road from Casamayo to Cajamarca, km 139 about 143 km southeast of Cajamarca, 2215 m, 7°17'08"S, 78°30'91"W, S. Stern, E. Tepe, S. Leiva & M. Zapata 109 (BM [BM001016880], HAO†, NY [NY00986682], USM, UT). La Libertad: Prov. Otuzco, alrededores de Huaranchal, 2140 m, [7°41'21"S, 78°26'51"W], 6 February 1999, A. Sagástegui, S. Leiva & V. Quipuscoa 11608 (BM [BM000935134]).
Solanum dillonii is superficially similar to the widespread Solanum riparium Ruiz & Pav. and to the more southerly Solanum conglobatum Dunal. It differs from Solanum riparium in its violet flowers, acute leaf bases and pubescence; both stem and leaf trichomes of Solanum dillonii are very long-stalked and multangulate, while those of Solanum riparium are sessile and tend to have more rays (tending to echinoid sensu
Solanum dillonii is another species endemic to the Amotape-Huancabamba phytogeographic zone (
Species nova Solano verecundo M. Nee similis, sed foliis coriaceis superne nitidibus, subtus valde pubescentibus, trichomatibus peltatis, differt.
Peru: Pasco: Prov. Oxapampa, Oxapampa-Villa Rica road, 7 km from road head, 2120 m, 10°36'S, 75°20'W, 4 January 1984, D.N. Smith & J. Albán 5558 (holotype: USM! [USM-123391]; isotypes: MO! [MO-5784802], NY! [NY00723838]).
Treelet to small tree, 2.5–9 (–18) m tall, branching in the upper part of the stems. Stems densely pubescent with persistent short-stalked peltate trichomes 0.2–0.4 mm in diameter, the rays 20–30, fused for almost their entire length, the midpoint absent, the trichome center dark reddish brown; new growth densely pubescent with peltate trichomes like those of the stems, drying pale beige. Bark of older stems pale brownish tan from the persistent trichomes. Sympodial units plurifoliate, the branching dichasial. Leaves simple, 6.5–16 cm long, 2–5 cm wide, narrowly elliptic, coriaceous, strongly discolorous, the upper surfaces glabrous and shiny, dark green when fresh, drying dark olive green, the lower surfaces densely pubescent with short-stalked peltate trichomes to 0.5 mm in diameter with >20 rays, subtended by a dense layer of tangled sessile echinoid trichomes ca. 0.1 mm long, the lamina not visible; primary veins 16–20 pairs, deeply impressed above, densely covered by pubescence beneath; base acute; margins entire, revolute; apex acute to acuminate; petioles 0.6–2 cm long, densely pubescent with peltate trichomes like those of the stems and leaf undersurfaces. Inflorescences terminal, 8–15 cm long, many times branched, with 60+ flowers, densely pubescent with peltate trichomes like those of the stems; peduncle 3–6 cm long; pedicels 5–8 mm long, ca. 1.5 mm in diameter at the base, ca.2 mm in diameter at the apex, stout, nodding at anthesis, densely pubescent like the inflorescence axes, articulated at the base; pedicel scars closely and more or less regularly spaced ca. 1 mm apart. Buds globose, the corolla exserted about halfway from the calyx tube just before anthesis. Flowers all perfect, 5-merous. Calyx tube 1.5–2 mm long, cup-shaped, narrowing gradually to the pedicel, the lobes 1.5–2 mm long, deltate, densely pubescent abaxially with peltate trichomes like those of the inflorescence rhachis, the adaxial surface sparsely pubescent with sessile echinoid trichomes. Corolla 1.2–1.5 cm in diameter, white, stellate, lobed ca. ¾ of the way to the base, the lobes 6–7 mm long, 4–4.5 mm wide, reflexed at anthesis, the tips and margins densely pubescent on the abaxial surface with peltate trichomes with >20 rays like those of the inflorescence, the adaxial surface glabrous, the tips and margins with a few sessile echinoid trichomes. Filament tube 0.5–1 mm long, the free portion of the filaments 1–1.5 mm long, glabrous, with tiny projections ca. 0.5 mm long on edge of tube between each filament; anthers 3–4 mm long, 1–1.5 mm wide, ellipsoidal, loosely connivent, yellow, poricidal at the tips, the pores lengthening to slits with age. Ovary densely pubescent with multangulate to echinoid trichomes; style 9–9.5 mm long, densely pubescent along its entire length with multangulate to echinoid trichomes with 4–30 rays ca. 0.2 mm long; stigma capitate, the surface minutely papillose, bright green in fresh plants. Fruit a globose berry, 0.6–0.7 cm in diameter, green when ripe, the pericarp thin, not shiny, unevenly pubescent with multangulate or echinoid trichomes with rays of many varying lengths, appearing scurfy; fruiting pedicels 1–1.1 cm long, ca. 1.5 mm in diameter at the base, woody, more or less erect. Seeds 40–50 per berry, 0.9–1 mm long, 0.9–1 mm wide, flattened-reniform, reddish or golden brown, the surfaces minutely pitted, the testal cells elongate.

Figure 4. Isotype specimen of Solanum oxapampense S. Knapp. (Smith & Albán 5558 NY [NY00723838]). Specimen image reproduced with the permission of The C. V. Starr Virtual Herbarium of The New York Botanical Garden (http://sciweb.nybg.org/science2/VirtualHerbarium.asp).
Endemic to the valley of the Río Huancabamba (Río Pozuzo) in central Peru; found within the Parque Nacional Yanachaga-Chemillen (Fig. 2).
Montane forests from 1500 to 2500 m elevation; as with many members of the Brevantherum group, often found along roadsides and in open areas.
Named for the valley of Oxapampa, where this species is endemic.
Solanum oxapampense
is known from only 4 localities, has an EOO of 198 km2 and AOO (using a
cell size of 0.04) of 195 km2; giving it a status of Endangered (
Peru: Pasco: Prov. Oxapampa, Dist. Huancabamba, camino a Pozuzo, 1200–1400 m, 10°04.02S, 75°32.59W, 2 June 2004, R. Rojas, M. Huaman, A. Peña & J. Mateo 2490 (MO, NY [NY00824860], USM); Prov. Oxapampa, Dist. Oxapampa, Parque Nacional Yanachaga-Chemillen, Sector Chacos, 2471, 10°37'S, 75°17'W, 24 January 2004, R. Vasquez, A. Monteagudo, A. Peña & R. Francis 28939 (MO, NY[NY00829065], USM); Prov. Oxapampa, Dist. Oxapampa, Oxapampa-Villa Rica road, 2300 m, 10°39.59S, 75°19.33W, 22 November 2007, E. Tepe, S. Stern, J. Mateo, M.P. Moreno, R. Rojas 2227 (BM [BM001017349], USM, UT).
Solanum oxapampense is a very distinctive species with its coriaceous and strongly discolorous leaves, with the upper surfaces dark green and shiny and the veins deeply impressed, and the lower surfaces densely pubescent with pale tan peltate trichomes. It is similar to Solanum verecundum M. Nee with which it is broadly sympatric, but does not apparently co-occur. Solanum verecundum is found along the eastern slopes of the Andes from Ecuador to southern Peru, Solanum oxapampense is possibly derived from it. The two species share trichomes that are peltate to some degree, small white flowers and small fruits (orange in Solanum verecundum, apparently green at maturity in Solanum oxapampense). Solanum oxapampense differs from Solanum verecundum in its coriaceous, narrower leaves and in its pubescence, which is composed of truly peltate trichomes whose rays are fused for more than half their length and that lack midpoints. Trichomes of Solanum verecundum have rays that are only fused in the central half, near the midpoint; midpoints are always present, even if sometimes very small and nub-like.
In the original description of Solanum verecundum (
Neither Solanum oxapampense nor Solanum verecundum would have been included in section Brevantherum by
Ecuador: Sucumbios: El Salado, colecciones en el sendero a la finca del Sr. Segundo Pacheco, 1400 m, 13 October 1990, J. Jaramillo, E. Grijalva & M. Grijalva 13285 (holotype: QCA; isotype: NY! [NY00381798]).
Shrub to small tree, 4–14 m tall. Stems densely pubescent with persistent short-stalked porrect –stellate to somewhat peltate trichomes 0.2–0.3 mm in diameter, the rays 10–12, fused for less than half their length, the midpoint sometimes a short stub to 0.5 mm long; new growth densely pubescent with porrect-stellate trichomes like those of the stems, these drying pale golden-brown. Bark of older stems reddish gold from the persistent trichomes. Sympodial units plurifoliate, the branching dichasial. Leaves simple, 6–19 cm long, 2–10 cm wide, elliptic or narrowly elliptic, membranous or chartaceous, discolorous (“silvery beneath" fide Bohs 3361), the upper surfaces moderately and evenly pubescent with sessile and short-stalked porrect-stellate trichomes with up to 15 rays, the rays fused only in their lower part near the midpoint, the midpoint to 0.2 mm long, the lamina visible, the lower surfaces densely pubescent with short-stalked porrect-stellate trichomes to 0.4 mm in diameter with up to 16 rays, the rays fused only in the center, the midpoint to 0.05 mm long, the lamina not visible; primary veins 12–15 pairs, not markedly impressed above, densely covered by pubescence beneath; base acute to somewhat attenuate onto the petiole; margins entire, plane; apex acute to acuminate; petioles 1–1–3(–4) cm long, densely pubescent with porrect-stellate to peltate trichomes like those of the stems. Inflorescences terminal, 7–10 cm long, many times branched, with 100+ flowers, densely pubescent with porrect-stellate to peltate trichomes like those of the stems; peduncle 2–5 cm long; pedicels 5–6 mm long, 1–1.5 mm in diameter at the base, ca.1.5 mm in diameter at the apex, stout, nodding at anthesis, densely pubescent like the inflorescence axes, articulated at the base; pedicel scars closely and more or less regularly spaced ca. 1 mm apart. Buds globose, the corolla strongly exserted from the calyx tube just before anthesis. Flowers all perfect, 5-merous. Calyx tube 1–1.5 mm long, cup-shaped, narrowing gradually to the pedicel, the lobes 1–1.5 mm long, deltate, abaxially densely pubescent with porrect-stellate to slightly peltate trichomes like those of the inflorescence, the adaxial surface sparsely pubescent with sessile porrect-stellate trichomes. Corolla 1–1.2 cm in diameter, white, stellate, lobed ca. ¾ of the way to the base, the lobes 4–5 mm long, 2–2.5 mm wide, reflexed at anthesis, the tips and margins densely pubescent on the abaxial surface with porrect-stellate trichomes with ca. 10 rays like those of the inflorescence, the adaxial surface glabrous, the tips and margins with a few sessile porrect-stellate trichomes. Filament tube minute, the free portion of the filaments ca. 1 mm long, glabrous; anthers 2.5–3 mm long, ca. 1 mm wide, ellipsoidal, loosely connivent, yellow, poricidal at the tips, the pores lengthening to slits with age. Ovary densely pubescent with multangulate to porrect-stellate trichomes; style 6–6.5 mm long, densely pubescent along its entire length with porrect-stellate 4–6-rayed trichomes ca. 0.2 mm long, the midpoints elongate and equal to the rays; stigma capitate, the surface minutely papillose. Fruit a globose berry, 0.5–1 cm in diameter, bright orange when ripe, the pericarp thin, not shiny, unevenly pubescent with multangulate trichomes with rays of many varying lengths, appearing scurfy; fruiting pedicels 0.9–1 cm long, 1.5–2 mm in diameter at the base, woody, erect. Seeds >100 per berry, 1–1.5 mm long, 1–1.5 mm wide, flattened-reniform, pale golden-yellow, the surfaces minutely pitted, the testal cells square.

Figure 5. Isotype specimen of Solanum verecundum M.Nee. (Jaramillo et al. 13258, NY [NY00381798]). Specimen image reproduced with the permission of The C. V. Starr Virtual Herbarium of The New York Botanical Garden (http://sciweb.nybg.org/science2/VirtualHerbarium.asp).
Along the eastern slopes of the Andes from northern Ecuador to southern Peru (to the Department of Cuzco; see Nee, 2000 for a distribution map).
Solanum verecundum occurs in premontane and montane forests, primarily in secondary growth (“purma") and along roads, from 1200–2000 m.
Solanum verecundum
is a relatively common species with a broad distribution along the
eastern slopes of the Andes and can be considered of Least Concern (
Ecuador: Napo: road Baeza-Tena, 8 km from Baeza, towards Río Cosanga, 1800–1900 m, 0°31'S, 77°50'W, 28 October 1976, H. Balslev & E. Madsen 10400 (BM [BM000935121]). Zamora-Chinchipe: road between El Progreso and Guaramizal, ca. halfway to Guaramizal, 1430 m, 4°48.23S, 79°07.26W, 28 March 2005, L. Bohs, J.L. Clark, J.R. Bennett, N. León et al. 3324 (BM [BM000846206]); Cantón Chinchipe, Parroquia Zumba, trail from Guaramizal to cabin of Sandy León, W of Escuela Byron Jiménez, just S of Las Pircas, 1600 m, 4°46.21S, 79°10.36W, 30 March 2005, L. Bohs, J.L. Clark, J.R. Bennett, N. León et al. 3361 (BM [BM000846204]); Cantón Valladolid, Parroquia Valladolid, road between Valladolid and El Porvenir del Carmen, close to Tapala, 1600–1680 m, 4°32.45S, 79°06.08W, 1 April 2005, L. Bohs, J.L. Clark, J.R. Bennett &N. León 3381 (BM [BM000846226]); Palanda, region de la Cordillera del Condor, Parroquia San Francisco de Vergel, riberas del Río Vergel, entre Santa Rosa y La Canela, 1200 m, 4°39.07S, 79°01.41W, 6 March 2007, W. Quizhpe & A. Wisum 2492 (BM [BM000943415]). Peru. Cajamarca: Prov. San Ignacio, San José de Lourdes, entre Camaná y Santo Tomás, 1800–1870 m, 5°01.00S, 78°52'W, 8 April 1997, J. Campos & S. Corrales 3801 (BM [BM000846199]); Prov. San Ignacio, San José de Lourdes, caserio Rumichina, 1679 m, 5°49.09S, 78°17.04W, 30 June 2006, J. Perea & V. Flores 2508 (BM [BM000943431]). Huánuco: Prov. Ambo, 38 km from Tingo Maria, between Tingo Maria and Pucallpa, 1680 m, 4 August 1978, J. Aronson & P. Berry 618 (BM [BM000795484]).
As mentioned above, the original description of Solanum verecundum (
I thank the C. V. Starr Virtual Herbarium of The New York Botanical Garden (http://sciweb.nybg.org/science2/VirtualHerbarium.asp) for permission to use images of specimens in their care; Kim Watson (NY) for scanning images of NY sheets; Guy Atchison (BM) for the map and for performing the conservation assessments; Michael Nee (NY) and Lynn Bohs (UT) for many long discussions about Solanum taxonomy; Nroman Robson (BM) for help with Latin; Eric Rodriguez (HUT), Asunción Cano and Maria Isabel Torre (both USM) for checking the holdings of herbaria in Peru; and the curators of the herbaria mentioned in the text for permission to examine specimens in their care; this work was funded by the National Science Foundation's Planetary Biodiversity Inventory (PBI) Program (DEB–0316614).