





(C) 2012 Peter H. Hovenkamp. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The Asiatic species of Oleandra (Oleandraceae) are revised. We reduce a large number of species to Oleandra neriiformis and Oleandra sibbaldii, we provide a revised circumscription of Oleandra cumingii and Oleandra undulata and we establish the identity of Oleandra vulpina. In total, we recognize 9 species, with full synonymy, descriptions and distribution maps. A list of identifications is appended.
Oleandra, systematics
Virtually all authors who have dealt with the genus Oleandra Cav. have commented on its distinctness or naturalness. The shrubby growth form, particularly distinct in Oleandra neriiformis Cav., prompted
Two genera have been described that are now universally included in Oleandra.
The characters of Oleandra were summarized most recently by
There is as much unanimity on the distinctness of the genus as there is uncertainty on the distinctness of species in Oleandra, and many authors who have dealt with the genus have commented on the difficulties of species delimitation.
Much of the variability between species in Oleandra is found in the rhizome, but as large parts of the rhizome are usually not preserved in herbarium collections, it is difficult to appreciate and describe this variability without field study. In the Asiatic species that we have seen, rhizomes may be relatively short-creeping, leading to more or less compact clumps of fronds, or more widely creeping. In the latter case, in some species the rhizome appears to be short-lived, decaying less than 1 m behind each growing point, which leads to stands with a scattered growth of plants. In others, the rhizome can be highly persistent, and stands may be extensive, with a dense growth of stems. In plants with this growth form, parts of the rhizome may also grow outwards or upwards from the substrate and form erect or drooping stems, here to be called “aerial” stems, that are often rootless. Branches are often opposite each other, and then both branches grow in the same direction, either downwards (positively geotropic) or upwards (negatively geotropic). Negatively geotropic branches can also be found on parts of the main rhizome where this starts to curve upwards, forming props that support the main stem. This is the shrubby growth form that is most often associated with the genus, although it occurs only in a minority of the species.
The anatomy shows a variably, but often very strongly developed peripheral sclerified sheath, a ground tissue with or without scattered sclerenchyma strands, and a dictyostele. When aerial parts develop, they tend to be more strongly sclerified.
The rhizome is covered with, usually persistent, rhizome scales with a peltate attachment. The scales are often strongly thickened near the attachment, and the margin can be nearly entire or densely set with woolly hairs (best visible in young scales) or sessile glands. The scales are strongly appressed or spreading to recurved - in the latter case the recurved parts tend to disappear on older rhizomes, giving the impression of a cover of short, appressed scales.
Roots arise mostly from the ventral side of the rhizome, and may be unbranched for a considerable length. The long unbranched parts have been described as rhizophores, but they were identified as real roots by
Fronds arise on the rhizome without any apparent regularity, sometimes clustered, sometimes more regularly spread over the length of a creeping rhizome. They do not appear to grow in regular rows, but are inserted more or less dorsally on creeping stems, and often on all sides on aerial stems. At a variable position on the stipe, there is a distinct articulation point, where old fronds abscise cleanly, with a plane of dehiscence that is perpendicular to the stipe. The part below the abscission point might be called a phyllopodium, the part above it the stipe, but the upper part of the phyllopodium is in all structural details similar to the stipe, while basally it shows a gradual transition to the rhizome. A stipe-like phyllopodium like this is restricted, in ferns, to Oleandra and Arthropteris J. Sm., but in the latter genus, the articulation is distinctly much more oblique. In other ferns where fronds are articulated to the rhizome (Polypodiaceae, Davalliaceae), the part below the articulation is more clearly rhizomatous in structure and indument. In the most inclusive phylogenetic analyses available (
The lamina is uniformly simple in all species, and varies little in shape, except in Oleandra werneri Rosenst.¸ which is dimorphic, with the fertile fronds or parts of fronds strongly contracted. Sori are indusiate with a reniform indusium, and always separate. The indusia may be firm and persistent or shriveling and inconspicuous in older sori. The sori are borne dorsally and singly on the veins, at distances from the costa that vary strongly. Sporangia are long-stalked, often with a number of glandular hairs attached to the distal part of the stalk, just below the capsule. The capsule is of the common Polypodiales type.
SporesFigs 1–10
The morphology of the spores of Oleandra as shown with SEM has been illustrated by
Two distinct types of perispore morphology can be distinguished. The first type occurs in all species studied except Oleandra wallichii. It is highly variable but the variation can be described in terms of just three parameters describing the folds, the ornamentation of the surface and the degree of perforation. This type shows a surface with a pattern of coarse folds, a variable ornamentation and a variable degree of perforation (“lophate”,
The second type occurs exclusively in Oleandra wallichii (figs 5, 6, 8). Here the perispore is composed of a thicker basal layer and thicker, more conical spines (fig. 8) with a basal diameter of 1–2 µm, which are, mainly in the specimens from Taiwan, partly fused in irregular ridges (fig. 7). In this type there never is an outer layer overlaying this pattern of conical spines, and thus the entire perispore is massive.
The exospore is smooth in all cases where it has been observed. Spore size variability was assessed based on SEM observations, with length of the spore measured including the perispore (Table 1, fig. 9). The limited data available do not allow a full statistical analysis, but it is clear that several species and even several specimens show a large variability in spore size. A similar variability in Oleandra from Southern Africa was found by
List of specimens (all L.) of which spores were studied (see figs. 9, 10).<br/>
| Specimen | Identification | number of spore measurements |
| Kato et al. B 9511 | Oleandra coriacea | 2 |
| Geesink & Santisuk 5384 | Oleandra cumingii | 3 |
| Schmutz 6086 | Oleandra cumingii | 3 |
| Ting & Shih 796 | Oleandra cumingii | 1 |
| Davidse & Sumithraarachchi 7965 | Oleandra musifolia | 5 |
| Holstvoogd 472 | Oleandra musifolia | 6 |
| Brooks | Oleandra musifolia | 4 |
| Kato, M. et al. C 4121 | Oleandra neriiformis | 2 |
| Kato, M. et al. C 1365 | Oleandra neriiformis | 4 |
| Clunie et al. LAE 63399 | Oleandra neriiformis | 4 |
| Degener 14279 | Oleandra neriiformis | 3 |
| Siew 125 | Oleandra neriiformis | 3 |
| Gaerlan et al. PPI 13079 | Oleandra neriiformis | 5 |
| Kato et al. B 7901 | Oleandra neriiformis | 4 |
| Kato et al. 1160 | Oleandra neriiformis | 5 |
| Croft 66 | Oleandra neriiformis | 4 |
| Craven & Schodde 133 | Oleandra neriiformis | 4 |
| Brass 29706 | Oleandra sibbaldii | 6 |
| Elmer 11451 | Oleandra sibbaldii | 2 |
| Hennipman 5430 | Oleandra sibbaldii | 2 |
| Kato et al. C 7480 | Oleandra sibbaldii | 1 |
| Sledge 1790 | Oleandra sibbaldii | 1 |
| Hennipman 3334 | Oleandra undulata | 4 |
| Maxwell 74/907 | Oleandra undulata | 2 |
| Banoc 3 | Oleandra undulata | 3 |
| Van Steenis 20870 | Oleandra wallichii | 4 |
| De Haas 2622 | Oleandra wallichii | 4 |
| Van Royen & Sleumer 5959 | Oleandra werneri | 5 |
| Braithwaite RSS 4045 | Oleandra werneri | 2 |

1 Oleandra cumingii, perispore with broad folds, low warty ornamentation, not perforated. scale bar 10 µm. Schmutz 6086 (L) 2 Oleandra cumingii, perispore with narrow wings, spiny ornamentation, few perforations. Scale bar 10 µm. Ting & Shih 796 (L) 3 Oleandra musifolia, perispore with wings partly replaced by fissures, irregularly warty ornamentation, few perforations. Scale bar 10 µm. Brooks s.n. (L 0317430) 4 Oleandra sibbaldii, perispore highly perforated. Scale bar 10 µm. Brass 29706 (L) 5 Oleandra wallichii, perispore spiny. Scale bar 10 µm. De Haas 2622 (L) 6 Oleandra wallichii, perispore spiny, spines confluent into ridges. Scale bar 10 µm. Van Steenis 20870 (L) 7 Oleandra sibbaldii. Perispore spines on a thin basal layer, exospore smooth, Scale bar 1 µm. Elmer 11451 (L) 8 Oleandra wallichii, perispore with thick basal layer, exospore smooth. Scale bar 1 µm. De Haas 2622 (L).
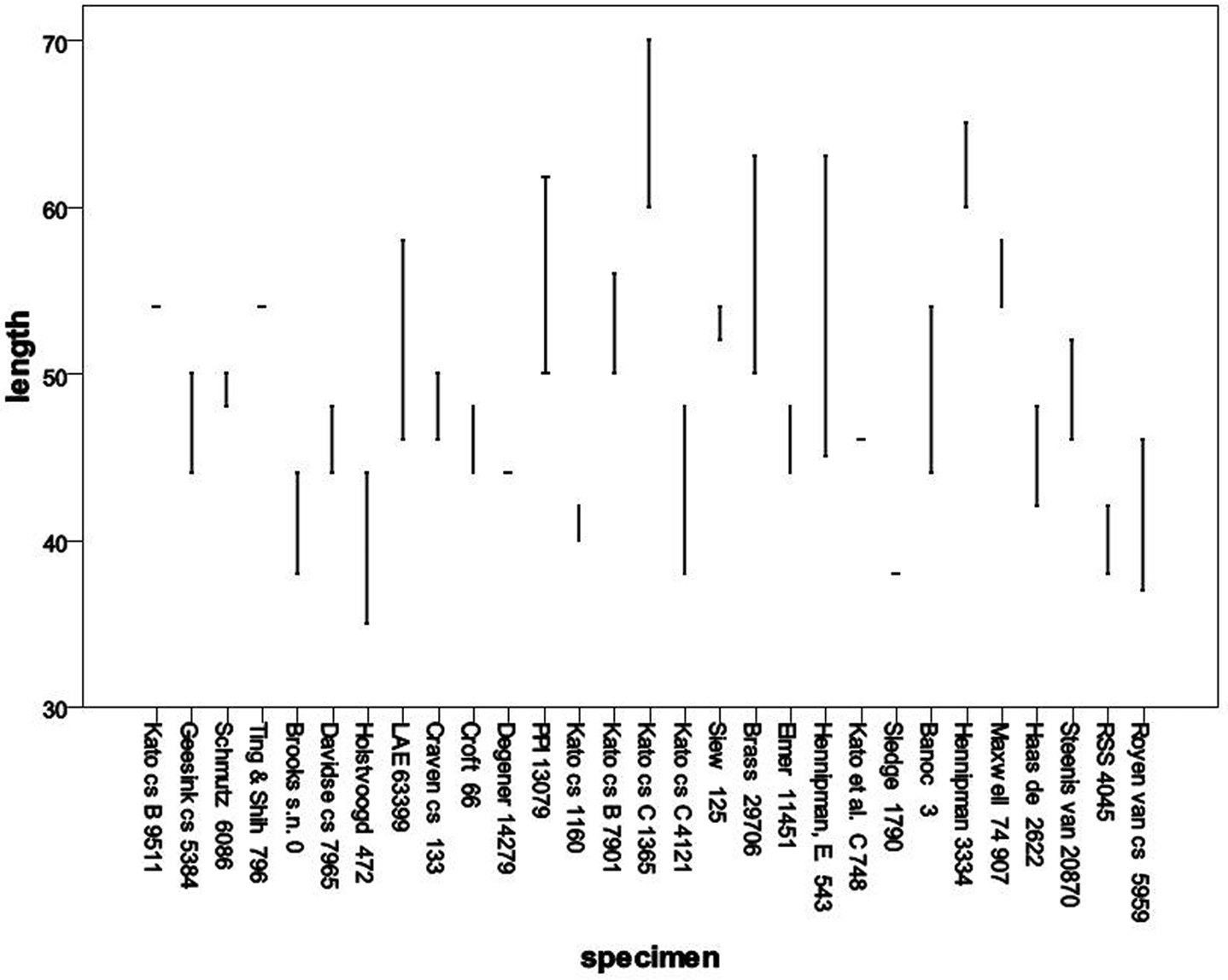
Distribution of spore lengths in Oleandra, arranged by specimen.
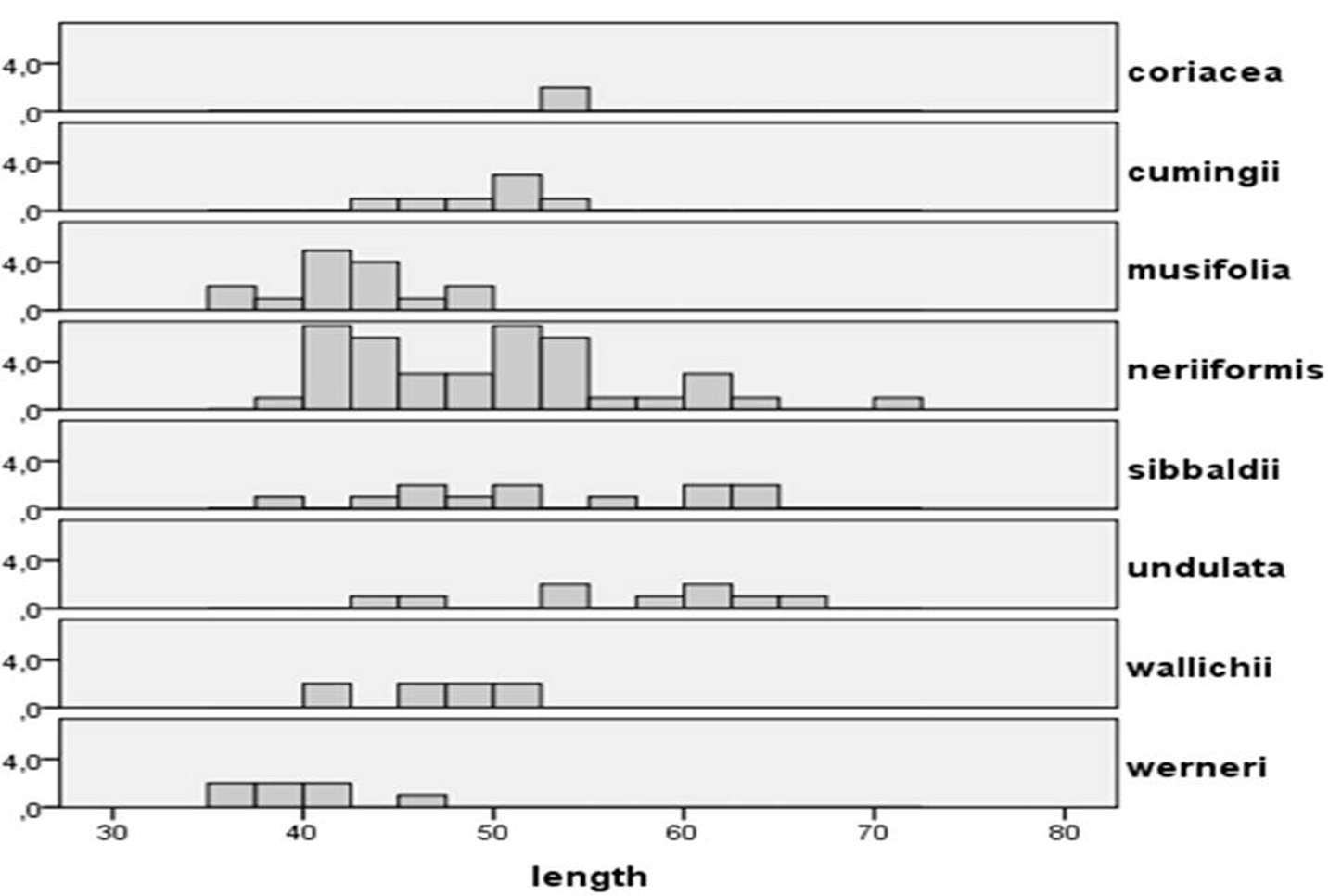
Distribution of spore lengths in Oleandra, arranged by species. Horizontal: spore length in µm, vertical: number of observed spores.
Data on chromosome numbers for species of Oleandra were summarized by
Of the Asiatic species, Oleandra neriiformis (1 count, source Himalayas), Oleandra musifolia (Bl.) C. Presl (2 counts, Ceylon and Southern India) and Oleandra wallichii (Hook.) C. Presl (2 counts, Northern India) have been reported. The report of a tetraploid taxon in Oleandra is based on a count for unsourced material growing in Kew (presumably from Africa), and is complemented by reports of n=41 for another plant from West Tropical Africa. The large variation in spore size in Oleandra from Southern Africa was conjectured by
The simple morphology of Oleandra has made comparison with other ferns difficult, and its position has been judged to be with Dryopteridaceae or Davalliaceae, where it has been associated in particular with Nephrolepis Schott and Arthropteris (
This study is based on material from B, BISH, BM, BO, K, KEP, KLU, L, P, PE, PNH, MICH, SING, UC (abbreviations follow Index Herbariorum, Thiers 2011). All specimens cited were seen except where noted otherwise. Specimens seen only as on-line images (provided either directly through the database of the holding institute or via JSTOR, http://plants.jstor.org) are marked with *. The identification list (see Appendix 1) has been prepared with the help of BRAHMS (http://dps.plants.ox.ac.uk/bol/BRAHMS), and the distribution data with BRAHMS and DIVA-GIS (http://www.diva-gis.org/).
http://species-id.net/wiki/Oleandra
Terrestrial, epilithic or epiphytic, creeping or scrambling ferns. Rhizome scaly, roots scattered, often with long rhizophore-like proximal parts, fronds scattered or in whorls, on stipe-like phyllopodia, dehiscing at a slightly thickened articulation point. Fronds stipitate, lamina simple, margin entire, veins distinct, somewhat raised on both sides, 1–2 ´ forked at or near the costa, costa often with narrow scales, lamina and veins often with acicular or capitate hairs. Sori in one, often irregular row on each side of the costa, with a more or less reniform, glabrous or hairy indusium. Sporangia stalked, stalk often with a number of sessile or stalked glands below the sporangium, sporangium body glabrous, spores monolete, perispore with broad wings, sometimes highly perforate, or echinate, massive.
| 1 | Rhizome with stiff, erect to pendent rootless aerial branches | 2 |
| – | Rhizome creeping | 4 |
| 2 | Fronds strongly dimorphic, scattered on the rhizome or somewhat clustered on short side branches | 9 Oleandra werneri |
| – | Fronds monomorphic or slightly dimorphic, often in whorls of 5–10 fronds | 3 |
| 3 | Lamina thick, coriaceous, costa with copious, conspicuous, 3–4 mm long pale to brown scales on the abaxial surface | 1 Oleandra coriacea |
| – | Lamina thin, papyraceous when dry, costa mostly with few or inconspicuous scales on the abaxial surface | 4 Oleandra neriiformis |
| 4 | Rhizome in older parts not entirely covered with scales; scales with squarrose acumen and entire or distinctly glandular margin | 5 |
| – | Rhizome entirely covered with overlapping scales; scales with appressed or spreading apex, usually with non-glandular cilia | 6 |
| 5 | Rhizome scales with gradually narrowed apex, stipes 0.5–4.5 cm long, costa and stipe often with distinct dark colouration on the abaxial surface | 5 Oleandra sibbaldii |
| – | Rhizome scales with abruptly narrowed apex, stipe to 2–3 mm long, costa and stipe without dark colouration on the abaxial surface | 7 Oleandra vulpina |
| 6 | Phyllopodia , inconspicuous, 2–5 mm, rarely 1 cm long, much shorter than the stipe, sori mostly in a closely costal single row | 7 |
| – | Phyllopodia, conspicuous, 2–10 cm long, often as long as or longer than the stipe, position of sori costal to medial | 8 |
| 7 | Rhizome scales squarrose, costa and stipe usually with dark colouration, frond apex distinctly apiculate | 8 Oleandra wallichii |
| – | Rhizome scales appressed, costa and stipe without dark colouration, frond apex acute to acuminate | 3 Oleandra musifolia |
| 8 | Rhizome scales spreading, long triangular with a wide acumen, brown, central part not conspicuously thickened, roots branching with root hairs over their entire length; lamina glabrous on upper surface and margin | 6 Oleandra undulata |
| – | Rhizome scales appressed to spreading, narrowly ovate-lanceolate with long narrow acumen, central part dark, thickened, roots often with glabrous, unbranched part (“rhizophore”), lamina hairy or glabrous | 2 Oleandra cumingii |
http://species-id.net/wiki/Oleandra_coriacea
Map 1, fig. 11 d–fRhizome with creeping parts unknown, aerial stems unbranched and rootless, 2–3 mm thick (when dry), not white waxy, in cross-section with a distinct sclerified peripheral sheath and few, scattered sclerified strands, phyllopodia in weak to distinct clusters of 4–6, short, usually less than 5 mm long. Scales persistently covering the rhizome, peltate, 5–70 × 0.5–1 mm, appressed (sometimes spreading), dark, shining with pale to brown margin and acumen, margin ciliate especially when young. Fronds monomorphic; stipe 1–1.5 cm long, without dark colouration, with up to 1 mm long glandular hairs and often small, appressed scales; lamina 13–30 × 1.2–3.3 cm, linear, base narrowly cuneate to rounded, apex acuminate to caudate with cauda to 1.5 cm, texture coriaceous; costa and veins on lower surface densely hairy with up to 1 mm long, acicular hairs, costa without dark colouration, with copious, conspicuous, 3–4 mm long pale to brown scales, upper surface more glabrous, mostly hairy on the costa only, with similar hairs and with less copious scales. Sori in a single more or less irregular medial row, separated from the costa by a 2–7 mm wide sterile zone, indusium distinct, c. 1.5 mm wide, most often glandular. Sporangial stalk with glands below the sporangium. Spores with coarse confluent ridges, areolae with short pointed excrescences, perispore hollow, with internal baculae, outer layer distinctly perforated.
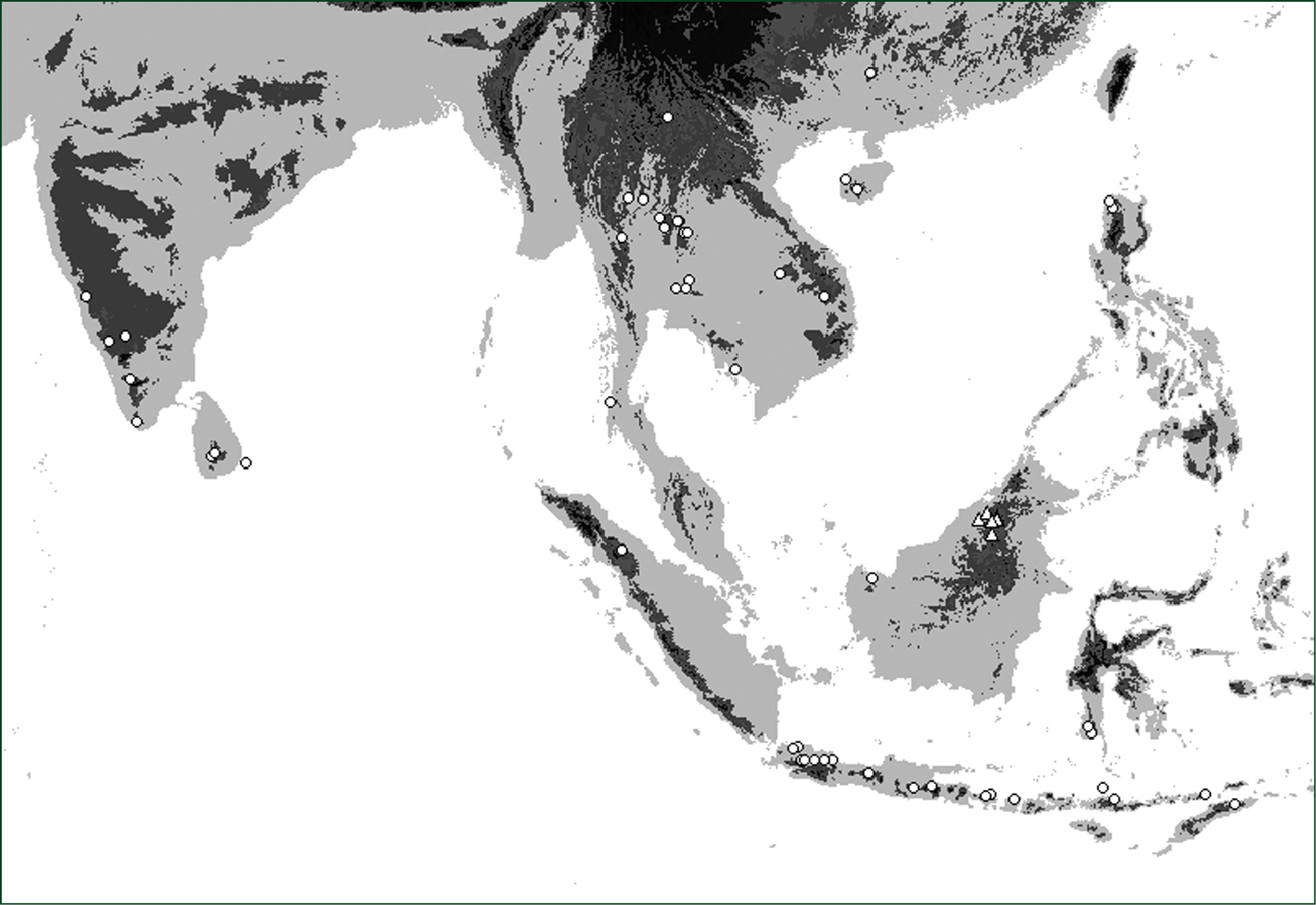
Distribution of Oleandra coriacea (triangles), Oleandra musifolia (circles).
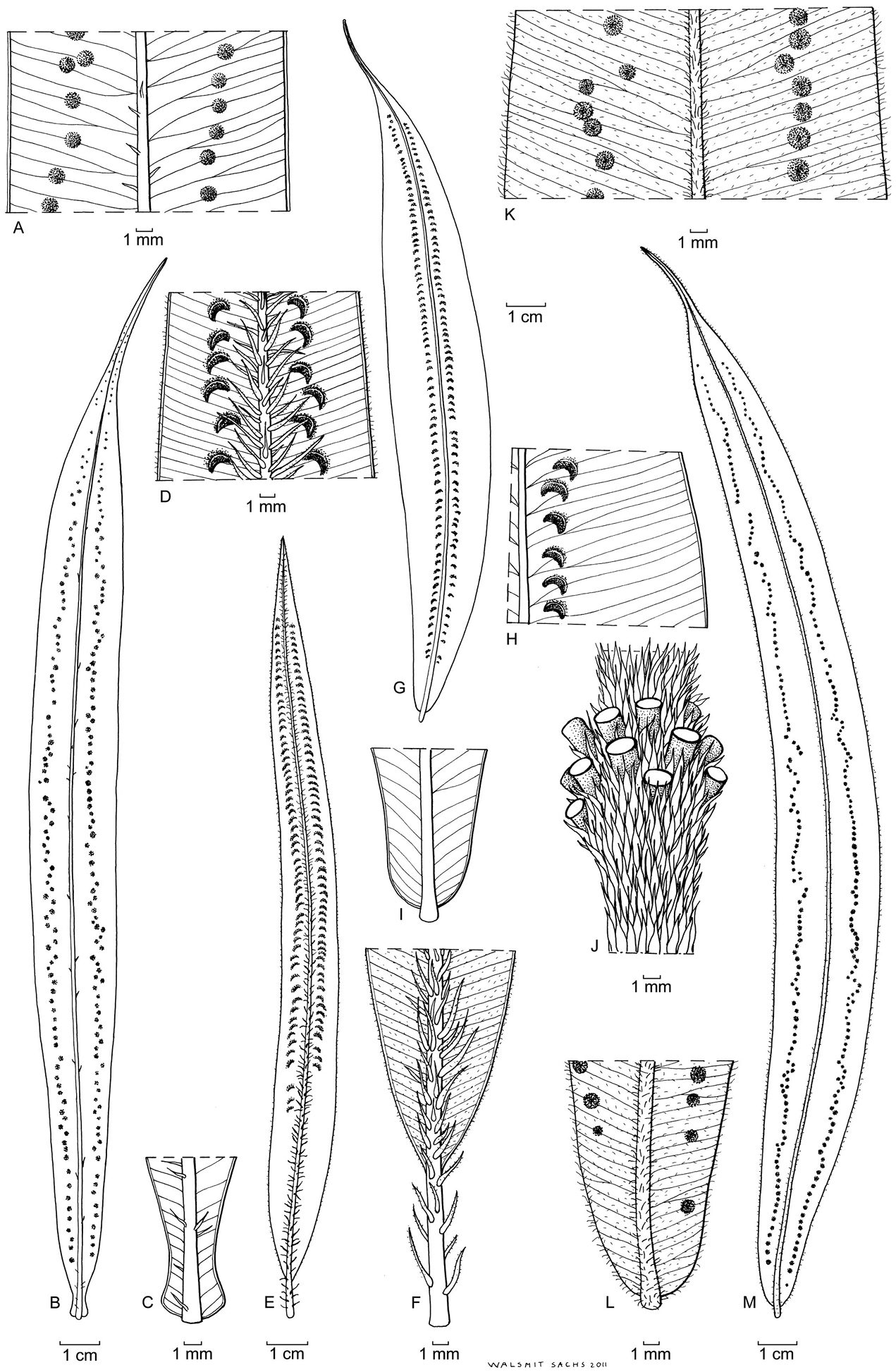
Oleandra neriiformis (a–c, g–m), Oleandra coriacea (d–f). a middle part of lamina, medial sori with indistinct indusia b entire lamina c lyrate lamina base with stipe articulation immediately below lamina base: Brass 23016 (New Guinea) d middle part of lamina, with conspicuous scales on midrib and sori with persistent indusia e entire lamina f lamina base with elongated stipe: Yahud et al SAR 88396 (Borneo) g entire lamina with costal sori h middle part of lamina, sori costal with persistent indusia i rounded lamina base with short stipe j whorl of short phyllopodia on aerial stem: Chew, Corner & Stainton 298 (Borneo) k middle part of lamina sori with inconspicuous indusium l rounded lamina base with short stipe m lamina: Copeland s.n. 29 jan 1933 (Philippines)., All views of lamina showing abaxial surface; all specimens L.
Brunei, Indonesia: Kalimantan Timur; Malaysia: Sarawak.
E
Terrestrial in montane forest, mainly on ridges and in summit vegetation, 1100–2200 m.
Creeping and rooting parts of the rhizome are absent in all collections seen, but presumably present, as in Oleandra neriiformis. The lamina of Oleandra coriacea is indeed much more coriaceous than Oleandra neriiformis, and has copious large pale costal scales and long hairs especially on lower surface. The sori are consistently medial, with firm indusia that are often clearly glandular on their surface.
http://species-id.net/wiki/Oleandra_cumingii
Map 2Rhizome short- to long creeping, 3–8 mm thick, sometimes white waxy in the older parts, little branched and not forming extensive stands, in cross-section with or without scattered sclerified strands; roots scattered, sometimes with unbranched aerial parts; phyllopodia scattered or more or less tufted, (1–)3–10 cm long. Scales persistently covering the rhizome, peltate, 4–9 × 0.5–1.5 mm, appressed, acumen with dark center and lighter acumen and margin, margin ciliate especially when young. Fronds monomorphic, stipe 2–12 cm long, without dark colouration, glabrous or hairy with catenate to acicular up to 2 mm long hairs, lamina 12- 40 × 2–4.5 cm, base narrowly cuneate to truncate, apex acute to long-acuminate, texture thin-chartaceous, both surfaces and margin with catenate or acicular hairs 0.2–1 mm long, usually more densely on lower surface, costa without dark colouration, on lower surface without or with few, pale to dark scales. Sori close to or scattered up to 2 mm from the costa, indusium distinct, 1–2 mm wide, densely hairy with short or long hairs. Sporangial stalk with glands below the sporangium. Spores with broad or narrow confluent ridges, surface variably ornamented with small pustules to narrow spines, perispore hollow, with internal baculae, outer layer not or finely perforated, sometimes fissured along the ridges.
China: Yunnan, Guangdong, Guangxi, Guizhou; Laos; Thailand (Peninsular); Malaysia: Peninsular Malaysia; Indonesia: Flores; Timor Leste; Philippines: Luzon.
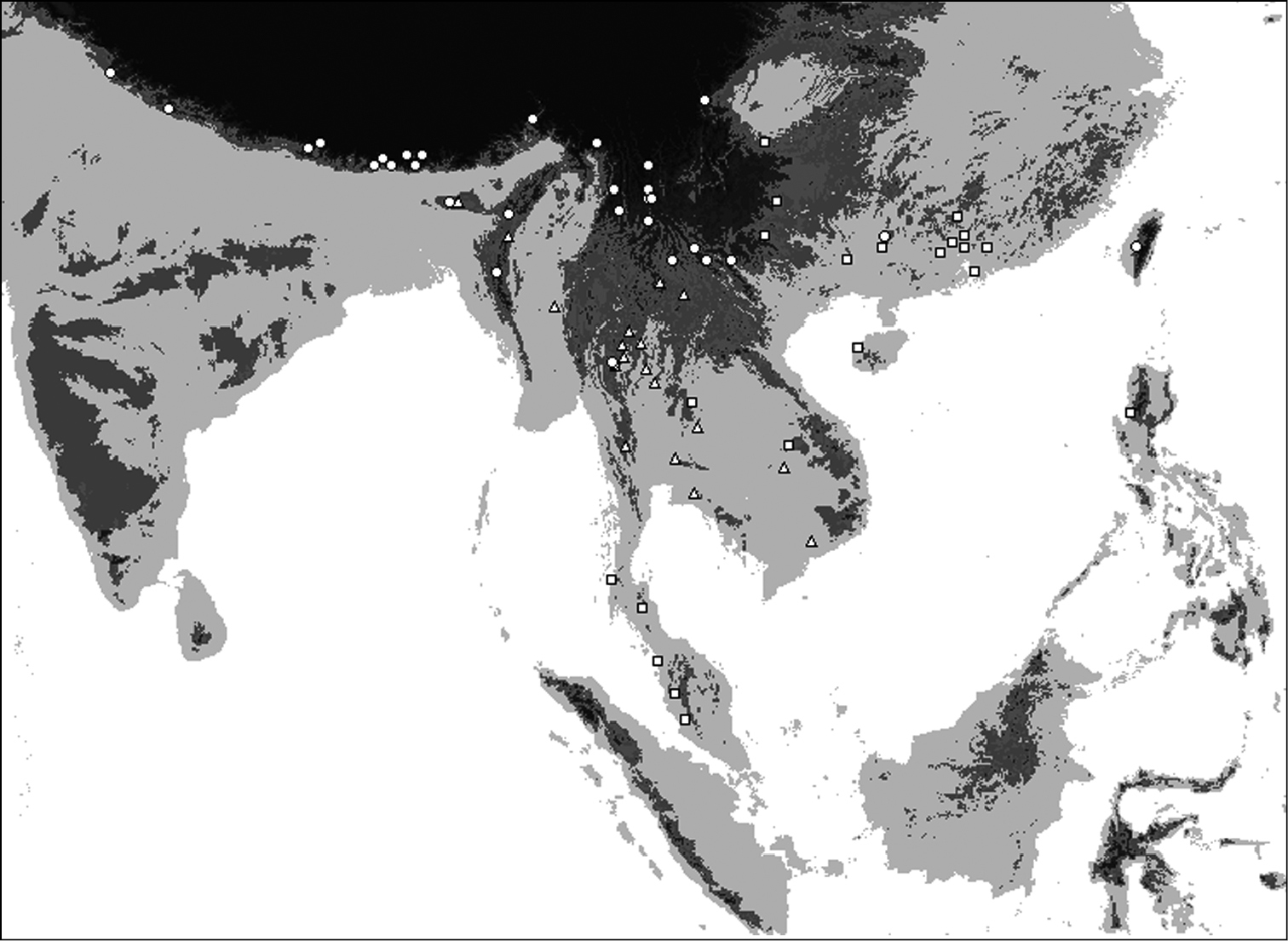
Distribution of Oleandra cumingii (squares), Oleandra undulata (triangles), Oleandra wallichii (circles).
Terrestrial or on rocks, cliffs and roadsides in open forest, sea level to c. 1200 m.
Contrary to
Oleandra cumingii is a variable assemblage of fairly widely scattered forms, in many characters intermediate between Oleandra undulata and Oleandra musifolia. From Oleandra undulata it can be distinguished by the more narrowly elongated, appressed rhizome scales with a more or less thickened costa in the acumen, and by the often hairy upper surface of the lamina. From Oleandra musifolia, Oleandra cumingii differs in the elongated phyllopodia and the distinctly tomentose indusia and lower surface of the lamina. The differences with either species, however, are often slight and bridged by specimens with an intermediate character combination. This may indicate that Oleandra cumingii is a hybrid or a hybrid swarm between the two other species, but failing positive evidence for that interpretation, we prefer to regard it here as a separate species.
http://species-id.net/wiki/Oleandra_musifolia
Map 1Rhizome creeping, 5–8 mm thick, white waxy in the older parts, often supported above the substrate by unbranched stilt-like roots, dorsiventrally flattened, (strongly compressed, 4–6 mm wide when dry), with up to 10 cm long, curved internodes (only occasionally straight and then often much longer) terminating in a cluster of a few short phyllopodia, usually less than 5 (–15) mm long, often hidden by the scales, rhizome often innovating just below this cluster, lateral branches usually basal on the internodes, in opposite pairs; all parts in cross-section without or with few sclerified strands, roots scattered, with long unbranched aerial parts. Scales persistently covering the rhizome, peltate, 5–10 × 1–1.5 mm, appressed, with dark center and lighter brown acumen and margin, margin ciliate especially when young and with sessile glands. Fronds monomorphic, stipe 0.5–4 cm long, without dark colouration, with short, glandular hairs; lamina to 60 × 4.2 cm, linear, base cuneate to truncate or more or less rounded, apex acute to long-acuminate, texture thin-chartaceous, both surfaces with catenate, often glandular hairs 0.2–0.5 mm long; costa without dark colouration, on lower surface with inconspicuous, 1–3 mm long brown scales. Sori mostly in a single regular row close to the costa, sometimes more scattered over a 2–5 mm wide zone close to the costa or at a distance of up to 3 mm, indusium distinct, 1.5–2 mm wide, glabrous or glandular, sometimes setose. Sporangial stalk with glands below the sporangium. Spores with coarse confluent ridges, areolae with short pointed excrescences, perispore hollow, with internal baculae, outer layer not or hardly perforated.
South China, Southern India, Sri Lanka, Thailand, Indonesia: Java, Lesser Sunda Islands, Sulawesi; Philippines: Luzon, Australia: Queensland.
Mostly terrestrial or on rocks, less often as low trunk epiphyte, in various types of forests, often disturbed, sea level to c. 2000 m.
ICN 60.8 specifies that the spelling of the original epithet “musaefolia” should be corrected to “musifolia”.
Aspidium moritzii is one of the two species distinguished by
Oleandra musifolia is variable in the density of hairs is variable, and the distance of the sori to the costa. It can be distinguished from Oleandra cumingii mainly by the short phyllopodia (longer in Oleandra cumingii). Incomplete collections are easily confused also with Oleandra neriiformis but can often be recognized by the flatter, softer rhizome without sclerenchyma strands (rhizome more rigid, rounded, with sclerenchyma strands in Oleandra neriiformis) and by the catenate hairs with capitate or glandular apex (more acicular, not capitate in Oleandra neriiformis). Distinguishing these two species on basis of juvenile material, however, is difficult, and often impossible.
http://species-id.net/wiki/Oleandra_neriiformis
Map 3, fig. 11 a–c, g–m, 12Rhizome with main stems creeping or ascending, 3–8 mm thick, white waxy in the older parts, creeping parts sparsely rooting, branches often in opposite pairs, ascending parts rootless, at base propped up by downwards directed branches, ultimately aerial, erect or pendent; branches single or in opposite pairs, mostly directly above a frond cluster, all parts in cross-section with a peripheral sclerified sheath and scattered sclerified strands, phyllopodia on creeping parts few, scattered, on aerial parts in more or less dense, often whorled clusters and branches, short to 15 mm long. Scales persistently covering the rhizome, peltate, 4–6.5 × 1–1.5 mm, appressed to squarrose, with dark center and lighter margin and acumen, margin ciliate. Fronds monomorphic or weakly dimorphic, stipe short to 3.5 cm long, without dark colouration; fertile lamina 12–43 × 0.5–4.5 cm, base gradually narrowed to narrowly truncate, then often somewhat lyrate, apex acuminate or to c. 2 cm caudate; sterile, if present, usually slightly shorter and wider, to 36 × 5.5 cm; texture thin-chartaceous, costa and lamina on lower surface without dark colouration, glabrous or with up to 2 mm long hairs, costa often with up to 2 mm pale to dark narrow scales. Sori in a single row close to the costa, or more scattered over a 2–5 mm wide zone close to or at a distance of up to 4 mm from the costa, indusium inconspicuous to distinct, to c 1.5 mm wide, glabrous to hairy. Sporangial stalk with glands below the sporangium. Spores with coarse confluent ridges, surface pustulose or with pointed excrescences, outer layer variably perforated.
India (Himalayas), China (Xizang), Malesian archipelago to Australia, Pacific Islands (Fiji, Samoa).
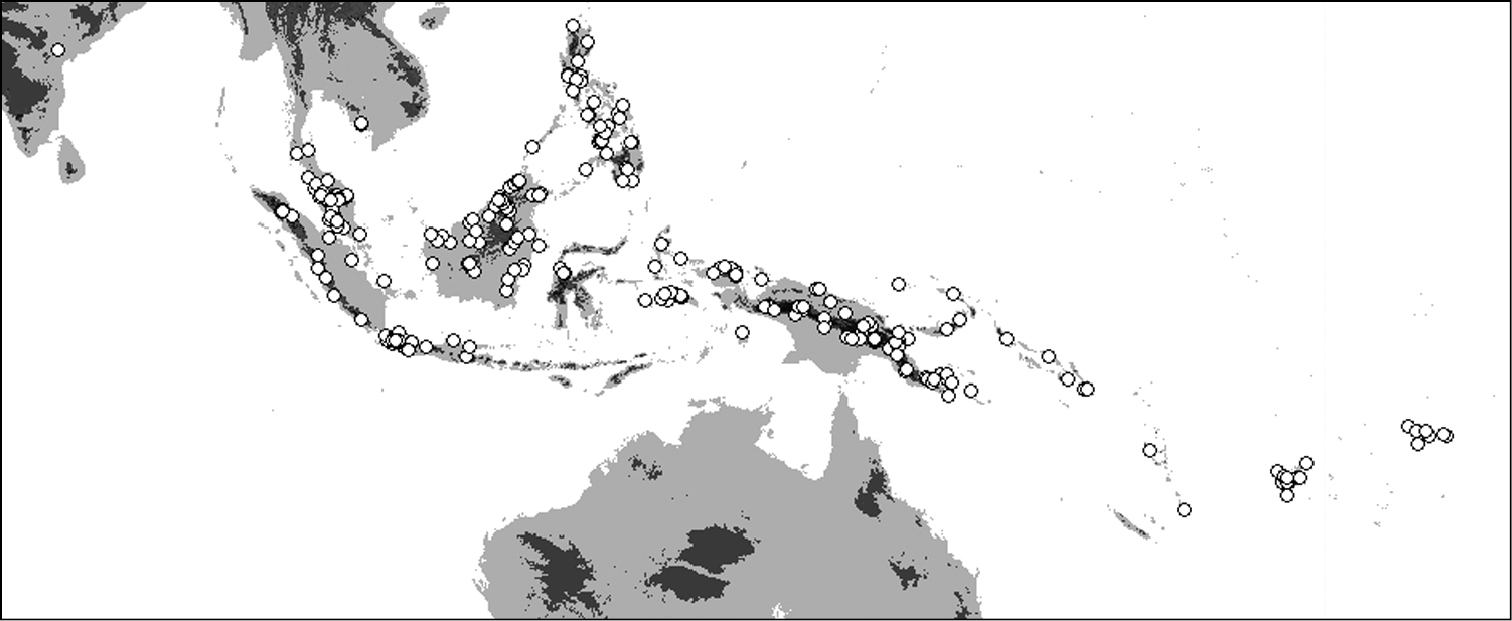
Distribution of Oleandra neriiformis.
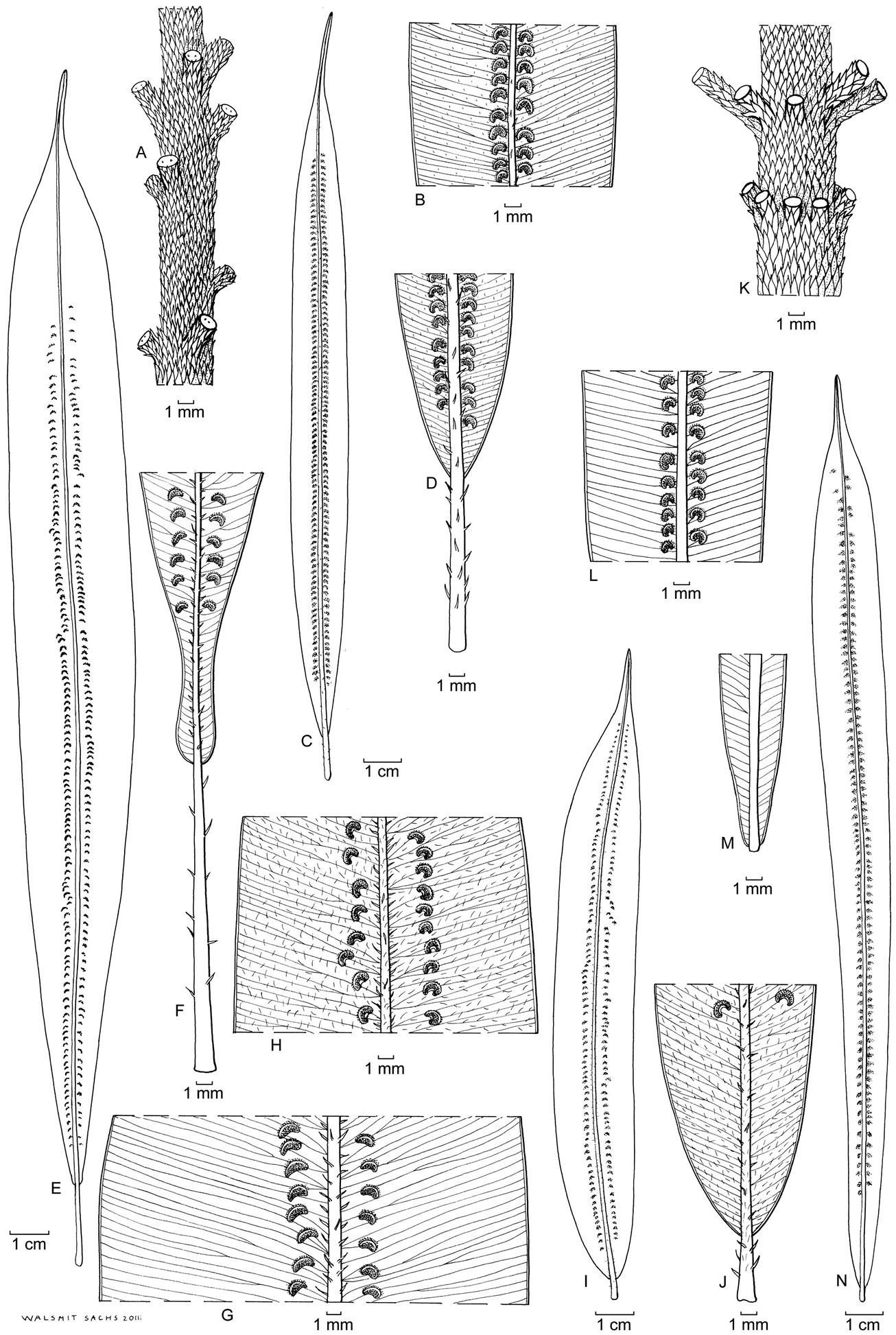
Oleandra neriiformis. a scattered phyllopodia on aerial stem b middle part of lamina with very closely costal sori and conspicuous indusia c lamina d lamina base with long stipe : A.C. Smith 6104 (Fiji) e lamina f lyrate lamina base with long stipe g middle part of lamina, sori costal with conspicuous indusia: Croft 66 (New Guinea) h middle part of lamina, sori costal with conspicuous indusia i lamina j cuneate lamina base with distinct stipe: Van Balgooy 5223 (Java) k irregularly whorled phyllopodia on aerial stem l middle part of lamina with very closely costal sori and conspicuous indusia m narrowed lamina base with articulation close to the lamina base n lamina: Chew W-L 942 (Peninsular Malaysia). All views of lamina showing abaxial surface; all specimens L.
Terrestrial or epiphytic, in various types of forests, in open places, often making up a significant part of summit or ridge scrub. Sea level to 2200 m.
The epithet neriformis published by Cavanilles has to be corrected to “neriiformis” (ICBN 60.8). The epithet neriifolia used by Presl is either a mistake to be corrected or a superfluous nom. nov. and should not be used.
Oleandra neriiformis does not occur in Australia. The reference in
Oleandra maquilingensis is based on two sterile specimens that are considered to represent juvenile plants by
Aspidium phyllarthron is one of the two species distinguished by
With its wide-creeping and persistent rhizome, Oleandra neriiformis may form extensive and probably long-lived stands, which, especially when it is collected a number of times over a long period, may give the impression of the presence of a locally abundant species with a highly constant and distinct combination of characters. We expect this is at least partly the basis for the multitude of local species that have been described, and that we all include in Oleandra neriiformis. Another problem is posed by the occurrence of juvenile plants in which the rhizome is not characteristically developed, and which may have much more softly hairy fronds than well-developed plants, blurring the distinctions to Oleandra musifolia and Oleandra cumingii.
The following characters or character complexes, some of which have been used to distinguish species, are variable in particular:
1 Place of the stipe articulation. The phyllopodium may be distinctly longer than the very short stipe (fig. 12 k, n), or a distinctly elongated stipe may be present equal to or longer than the phyllopodium (fig. 12 a, d)
2 Length and density of lamina hairs. Although the presence or length of lamina hairs is usually highly variable, some forms have constantly and distinctly longer hairs.
3 Location of soral zone. Sori may be located in a narrow zone close to the costa (fig. 12 b, n), or in a more irregular zone at some distance from the costa (fig. 11 b, m).
4 Indusium. The presence of an indusium is rarely constant over an area. It may vary from distinct and often firm (fig. 11 h, 12 b) to inconspicuous (then often hairy) or absent (fig. 11 a, k).
5 Costal scales: Some forms have uniformly pale and flat costal scales, some have almost uniformly narrow, dark scales, and there are forms that vary in this character.
Over most of the distribution area, two forms can often easily be distinguished locally, on basis of the relative length of phyllopodium and stipe. Stipitate forms have short phyllopodia, elongated stipes (thus the articulation is positioned at the base of the phyllopodium/stipe), the lamina gradually narrowed towards the base, and sori relatively close to the costa (fig. 12 k–m). This corresponds to the type of Oleandra neriiformis. The other form is characterized by longer phyllopodia, stipes short or absent (thus the lamina appearing sessile with regard to the articulation, fig. 2 b), usually a truncate lamina base (although the lamina directly above the base may be strongly narrowed, the base is still suddenly contracted, and often somewhat lyrate, fig. 2 c) that is clearly set off against the stipe, and sori in a more variable position, in some cases almost at the margin (fig. 2b). Other characters, such as indument, or indusium are independently variable and often vary in a similar way across the two forms where they co-occur, thus giving rise to different characteristic character combination in different parts of the distribution area.
Continental Asia. The few collections from continental Asia do not allow for an evaluation of the variability.
Java, Sumatra. On Java and Sumatra, there is no distinction between stipitate and sessile forms, as both stipe and phyllopodium lengths are strongly variable and extremes are not sharply separated. In
Peninsular Malaysia, Southern Thailand. In collections from the Malay Peninsula the difference between stipitate and sessile forms is associated with differences in hairiness of the lamina and position of the sori.
Borneo. On Borneo, the two forms also differ in hairiness of lamina, but not in the position of the sori, which is usually more or less closely costal. Here, the stipitate form is almost exclusively reported as epiphyte, the other form as terrestrial. Both forms tend to have narrower, darker costa-scales than in other areas. Completely glabrous forms such as are most common on Java and Sumatra are not found on Borneo. The sessile form has been described as Oleandra oblanceolata Copel. Stipitate form description: Phyllopodia up to 6 mm long, stipe 7–25 mm, lamina at base gradually narrowed, usually hairy with relatively long hairs (rarely glabrous), costal scales more or less frequent, mostly brown. Almost exclusively reported as epiphyte. Sessile form description (see also fig. 11 g–j): Phyllopodia 2–8 mm long, stipes at most 1 mm, lamina at base truncate, often ± lyrate, lamina mostly glabrous, sometimes short-hairy, costal scales not frequent, dark, narrow. Mostly reported as terrestrial.
Philippines. On the Philippines, distinctly stipitate forms represent a small minority of all collections (e.g., Cuming 94, Soejarto 8874, PNH 3862, 8710), but include the type of Oleandra neriiformis (
Copeland (1958) distinguishes Oleandra neriiformis from the short-stipitate form, and within the latter a number of species, based on details of indument: Oleandra herrei, with paleate costa, Oleandra colubrina, with setose costa and lamina, Oleandra nitida, with setose costa and glabrescent lamina. We find that although the density of costal scales varies strongly (it seems to be negatively and weakly correlated with the density of setae), scales can be found on all specimens, and the density of hairs on the lamina varies strongly. We see no basis on which these characters could lead to the distinction of clear groups.
Stipitate form description: Phyllopodia to 2 mm, stipes to c. 7 mm, lamina and margin hairy with highly variable density, costa mostly with many pale scales, sori not closely costal to medial or submarginal, indusia glabrous, often small, indistinct. Sessile form description (fig. 11 k–m): Phyllopodia 5–15 mm, stipes short, up to 1 mm long, otherwise similar.
Celebes, Moluccas. In eastern Malesia south of the Philippines, two forms co-occur on Celebes and the Moluccas, while the stipitate form extends to the Solomon islands and Vanuatu. Both forms here have indusia that are frequently shortly setose.
Stipitate form description: Phyllopodia 1–5 mm, stipes 8–25 mm, lamina base gradually narrowed, costal scales pale except near base of lamina, lamina setose, lamina hairs relatively long, margin often distinctly fimbriate with hairs shorter or equal to those on lamina, sori costal, indusia firm, with wide sinus, usually glabrous.
Sessile form description: Phyllopodia 5–15 mm, stipes very short to occasionally 3 mm long, lamina base narrowed to ultimately cuneate, not distinctly lyrate, lamina indument often long, conspicuous on all veins, margin often distinctly fimbriate with hairs similar to these on the lamina, costal scales few or absent, dark, sori narrowly costal, indusium distinct and persistent but not firm, often with narrow sinus, sometimes setose.
Kato (1989) distinguishes the specimens from Ceram with a very small, setose indusium as Oleandra cuspidata, but this represents only the extreme state of variability.
Fiji, Samoa. More eastwards in the Pacific Islands, there is no variability in the location of the articulation. All specimens from this area have elongated stipes, and can collectively be distinguished from the forms in other areas by the distinctly more lax clusters of fronds on the aerial stems (fig. 12 a), with the fronds mostly hardly clustered at all (a character in which they resemble Oleandra pilosa Hooker, from Tropical America). Regional differences show up mainly in the indument of the fronds. On Samoa, the lamina is less densely hairy than on Fiji, and the hairs on the lamina, when present, are distinctly shorter than those on the margin. Costa scales also differ, and are distinctly darker on Samoa than on Fiji, while sori are somewhat less strictly costal on Samoa.
Fiji form description (see also fig. 12 a–d): Phyllopodia 1–9 mm, stipes 5–35 mm, lamina base gradually narrowed, costal scales pale, lamina setose, lamina hairs 1–2 mm long, margin often distinctly fimbriate, hairs on margin shorter or at most equal to those on lamina, sori costal, indusia distinct, glabrous or setose,
Samoa form description: Phyllopodia 1–7 mm, stipes 2–28 mm, lamina base gradually narrowed, costal scales brown to dark, lamina glabrous or setose, with hairs to 0.5 mm long on the lamina, 1–2 mm long on the margin, sori costal or subcostal leaving a sterile zone of 0.5–3 mm wide, indusia distinct, glabrous or setose.
New Guinea and surrounding islands. The stipitate form that extends eastwards from Celebes co-occurs, on the main island of New Guinea, with a sessile form that is often distinctly dimorphic and has sori often quite distant from the costa.
Stipitate form description (see also fig. 12 e–g): Phyllopodia to 4 mm, stipes 5–25 mm, lamina base gradually narrowed, costal scales pale except near base of lamina, lamina setose, with hairs 0.5–1.5 mm long, margin glabrous, sori costal or subcostal leaving a sterile zone of 0.5–2.0 mm wide, indusia firm, with wide sinus, usually glabrous.
Sessile form description (see also fig. 11 a–c): Phyllopodia to 4 mm, stipes very short or absent, articulation directly below the base of the lamina, fronds slightly but usually distinctly dimorphic, lamina narrowed to a truncate or somewhat lyrate base, costal scales mostly brown to dark, lamina glabrous or sometimes short setose, with hairs to 0.5 mm long, margin usually with a few scattered pale, acicular hairs especially near the base, sori medial or submarginal leaving a sterile zone of 0.5–7 mm wide, indusia small and inconspicuous in older sori, occasionally more distinct, often fringed with pale acicular hairs.
http://species-id.net/wiki/Oleandra_sibbaldii
Map 4Rhizome long-creeping or pendulous, sparsely branching, 1.5–4.5 mm thick, very strongly white waxy, in cross-section without or with few scattered sclerified strands, phyllopodia scattered, (0.3–)0.8–4.0(–5.7) cm long, roots with distinct unbranched parts. Scales deciduous, exposing the rhizome in older parts, peltate, (3–)5–15(–18) × (0.3–)0.4–0.9(–1.2) mm, usually widest above the attachment, squarrose, reddish brown, margin entire or more or less densely set with sessile glands, acumen attenuated, long filiform apex. Fronds monomorphic; stipe (0.3–)0.5–3.5(–4.5) cm long, often with dark coloration on abaxial side; lamina to (4.5–)15–40(–58) × 1–4(–6) cm, widest in middle part, base attenuate to cuneate, sometimes rounded, rarely truncate, sometimes asymmetric, apex acuminate or up to 3 cm caudate, texture membranous to coriaceous, both sides sparsely to densely set with catenate hairs to c. 0.5 mm long; costa on lower surface often with dark-coloration, on both surfaces with scales, scales on lower surface often abundant, to 6(–11) × 1(–1.2) mm, pale to dark brown, on upper surface scarce, inconspicuous. Sori inframedial, leaving a distinct 1–4 mm wide sterile zone between costa and soral zone, sometimes as much as 12 mm from costa, indusium firm, to 1 mm wide, hairy. Sporangial stalk with glands below the sporangium. Spores with an irregular mesh-like network of up to 10 µm high folds, finely papillose with spinules up to ca. 4 × 1 μm, perispore baculate, outer layer much perforated.
Eastern Malesia to Pacific Islands. Malaysia: Sabah, Sarawak, Philippines: Mindanao; Indonesia: Sulawesi; Moluccas, Papua; Papua New Guinea; Solomon islands; Vanuatu; Tahiti and Marquesas: Hiva Oa, Nuku Hiva, Tahuata, Ua Huka and Ua Pou; Western Samoa: Savaii; Fiji.
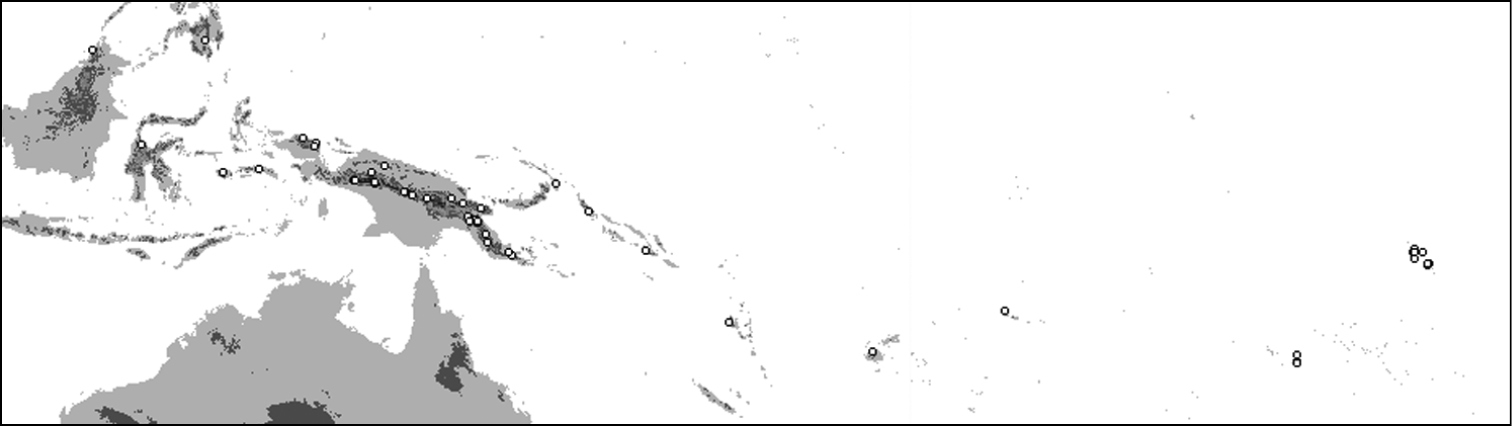
Distribution of Oleandra sibbaldii.
Epiphytic, epilithic, or less commonly terrestrial (most often at elevations over 1500m), commonly collected from mossy forests, climbing or sprawling among bryophytes and other epiphytes, or pendulous from mossy tree branches, to 600 m (Marquesas and Tahiti); or from 1000 to 3000 m (elsewhere).
Oleandra sibbaldii is common on New Guinea, but appears to be much sparser towards the periphery of the distribution area.
The dark colour on the abaxial surface of the costa is often very conspicuous, extending on the stipe, thus rendering the stipe conspicuously bicolorous.
http://species-id.net/wiki/Oleandra_undulata
Map 2Rhizome short-creeping, little branching, 5–6 mm thick (3–5 when dry), not white waxy, in cross-section with many scattered sclerified strands; phyllopodia irregularly scattered, close together or distant, 2–8 cm long, roots scattered, without distinct unbranched parts. Scales persistently covering the rhizome, peltate, 3–5 ×1–1.6 mm, slightly spreading, acumen brown, with sparsely ciliate margin, apex short, wide. Fronds monomorphic; stipe 3–15 cm long, without dark coloration, articulation at 1/5–1/2 from base; lamina to 60 × 1–5.5 cm, widest in middle part, base truncate or cuneate to gradually narrowed, apex acute to narrow-acuminate, texture herbaceous, upper surface glabrous or sparsely, lower surface more densely set with usually acicular, sometimes slightly catenate hairs to c. 0.5–1.0 mm long; costa on lower surface without dark coloration, without scales. Sori close to costa or leaving a distinct 1–6 mm wide sterile zone between costa and soral zone, indusium firm, to 2 mm wide, hairy. Sporangial stalk without glands below the sporangium. Spores finely papillose and coarsely ridged, perispore baculate, outer layer not perforated.
Burma, Laos, Thailand, China: Yunnan. Mostly below 1000 m.
In open or deciduous forests, often disturbed; terrestrial or epilithic, rhizome subterraneous, on rocks or in crevices, mostly on granite.
Oleandra undulata can be difficult to distinguish from Oleandra cumingii. In addition to the differences listed under that species, subterraneous growth of the rhizome may be characteristic for Oleandra undulata, but field observations are lacking for many specimens.
http://species-id.net/wiki/Oleandra_vulpina
Map 5Rhizome long-creeping, ca. 3 mm thick, not white waxy, sparsely branching, roots with unbranched parts; in cross-section with weakly developed sclerified shealth and few scattered sclerenchyma strands, phyllopodia scattered, 3–6 cm distant, 6–7 mm long. Scales scattered, not covering the rhizome, peltate, to 5 × 0.5 mm, appressed at the base, with a narrow squarrose acumen, dark brown near attachment, lighter towards margin, margin densely set with glands and multicellular hairs terminating in a gland. Fronds monomorphic; stipes with scales as the rhizome but less dense and with short fine glandular hairs; stipe 2–3 mm long, without dark coloration on abaxial side, bearing short fine glandular hairs; lamina linear-lanceolate, 17.5—20 × 3–4 cm wide, base cuneate, apex short caudate, tips up to 1 cm long, margin undulate, weakly cartilaginous, texture papyraceous, all parts with to 0.5 mm long catenate glandular hairs, or with longer, to 1.5 mm, acicular hairs; costa abaxially without dark coloration, with hairs like the stipe and with small scales; veins terminating in a weakly developed hydathode before the margin. Sporangial stalk with glands below the sporangium. Sori in an irregular row 2.5–6 mm from the costa, indusium round-reniform, c. 0.5 mm across, fugacious at very early stage. Spores absent.
New Guinea, at 975 m.
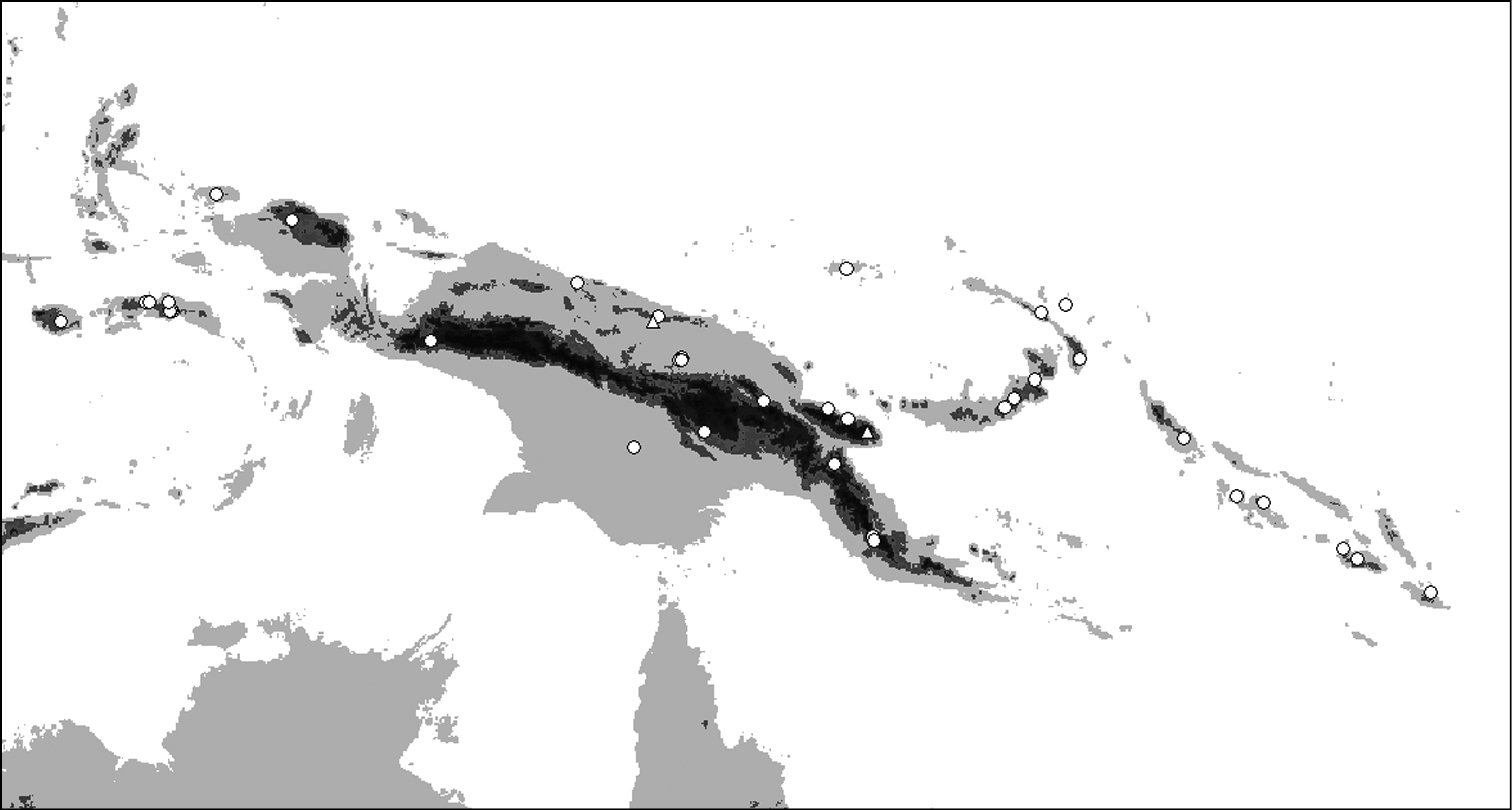
Distribution of Oleandra vulpina (triangles), Oleandra werneri (circles).
Scandent on trunk of Sago palm in garden, or in forest.
Taingelem (Wapi language, Miwaute)
Oleandra vulpina is distinct from Oleandra sibbaldii in the rhizome and costa scales. Rhizome scales are darker in colour with pale-coloured margin, long subulate apices, and margin strongly ciliate with glandular hairs. A similar glandular scale indument is found in the American Oleandra articulata (Sw). C. Presl, but not in the other Southeast Asian species. The costa scales beneath are small and inconspicuous, narrowly lanceolate. In addition, the rhizome of Oleandra vulpina is not glaucous. The indusia are very small and shrivel at a very young stage but are distinct when present and bear numerous setose hairs.
http://species-id.net/wiki/Oleandra_wallichii
Map 2Rhizome creeping, 3–4 mm thick (2–3 when dry), white waxy in the older parts, with long, leafless parts alternating with more or less dense clusters of very short phyllopodia, usually less than 2 (–5) mm high, of which usually only 1–2 bear fronds at the same time, branches usually in opposite pairs; in cross-section with weak sclerenchyma sheath and without sclerified strands, roots scattered, usually with long unbranched parts. Scales persistently covering the rhizome, peltate, 3–7 ×1–1.3 (–1.5) mm, base appressed, with dark center and lighter margin, acumen brown, usually recurved, with ciliate margin, apex twisted and with frizzly cilia. Fronds monomorphic; stipe 1–5 cm long, with dark coloration on abaxial side often distinctly bicolorous, with catenate hairs; lamina 13–45 × 2–4.5 cm, base truncate to rounded, apex often abruptly caudate, texture thin-herbaceous, translucent, both surfaces densely set with catenate hairs 0.5–1 mm long; costa on lower surface in basal half of lamina with dark coloration, with copious 2–4 mm long pale scales. Sori close to costa, indusium thin, to 1 mm wide, glabrous or hairy. Sporangial stalk without glands below the sporangium. Spores echinate and ridged, perispore solid.
Himalayas to Northern Thailand, Yunnan and Taiwan, 1600 to 3600 m. India, Nepal, Bhutan, China (Yunnan, Taiwan), Thailand.
Mostly epiphytic, on mossy trunks, also on cliff faces or boulders.
Oleandra wallichii differs from Oleandra undulata in the more wide-creeping rhizome that is often distinctly glaucous beneath and between the scales, without sclerenchyma strands; the often conspicuously bicolorous stipe, lamina with usually very distinctly apiculate apex and costa with frequent scales, sori constantly closely costal, with small indusia.
http://species-id.net/wiki/Oleandra_werneri
Map 5Rhizome with main stems creeping, scrambling or pendent, 2–3 mm thick, white waxy in the older parts, sparsely rooting, mostly terete when dry, bearing scattered short, usually less than 5 mm long phyllopodia, phyllopodia on aerial parts more closely together but not densely clustered; all parts in cross section with a peripheral sclerified sheath and scattered sclerified strands. Scales covering the rhizome, peltate, 3–4 × 0.5–1 mm, somewhat squarrose, with dark center and lighter margin and acumen, margin ciliate especially when young. Fronds strongly dimorphic, stipe 0.5–2.5 cm, without dark colouration, especially upwards with up to 2 mm long hairs; lamina chartaceous; fertile 27–56 × 0.5–1 (–1.7) cm, base and apex narrow, sterile 17–30 × 2.2–5.7 cm, base mostly cuneate, apex distinctly 2–3.5 cm caudate; costa and lamina on lower surface glabrous or with up to 2 mm long hairs, costa without dark colouration, like the stipe with up to 2 mm long brown scales. Sori in a single row on both sides of the costa, indusium firm, 1–3 mm wide, glabrous. Sporangial stalk with glands below the sporangium. Spores with coarse confluent ridges, areolae with short pointed excrescences, perispore hollow, with internal baculae, outer layer not or hardly perforated.
Indonesia (Maluku, Papua); Papua New Guinea; Vanuatu.
Commonly epiphytic, on trunks or in crowns, less often terrestrial or on rocks, erect, scrambling or pendent, in various types of forests, most frequently in montane or mossy forests, on ridges, up to c. 2000 m.
As in Oleandra neriiformis, rhizome morphology is probably more complicated than can be inferred from the mostly aerial unbranching parts making up most of the collected material.
Hairiness is very variable, and while there is no sharp distinction between hairy and glabrous forms, it is noteworthy that hairy forms tend to occur at especially the Western extreme of the distribution area, with less hairy forms near the Eastern extreme and glabrous forms mostly on the mainland of New Guinea.
We are grateful for the directors of the herbaria of BISH, BM, MICH, PNH, SING, UC for the loan of material, and of B, BM, BO, K, KEP, KLU, P, PE for the permission to study their collections. The contributions of many staff members in L, but in particular Peter van Welzen and Niels Raes, to the georeferencing of collecting localities and thus to the maps presented here are gratefully acknowledged.
Oleandra coriacea: 1
Oleandra cumingii: 2
Oleandra musifolia: 3
Oleandra neriiformis: 4
Oleandra sibbaldii: 5
Oleandra undulata: 6
Oleandra vulpina: 7
Oleandra wallichii: 8
Oleandra werneri: 9
Abbe, L.B.; Abbe, E.C. 9650 : 4; 9805 : 4; Abdul Samat, A. 118 : 4; Adelbert, A.G.L. 128 : 4; 252 : 4; 490 : 4; Aet; Idjan 324 : 4; Ajoeb 265 : 4; Allen, B.M. 1427 : 4; 1779 : 4; Alston, A.H.G. 16962 : 4; Anderson, J.A.R. S 18570 : 4; Andrews, S.B.; Stocker, G. 283 : 3; Ashton, P.S. 242 : 4; 424 : 1.
Backer, C.A. 12576 : 3; 23016 : 4; 25877 : 4; 36909 : 3; Bakhuizen v.d. Brink, R.C. 1539 : 4; 2777 : 4; 3208 : 4; 4482 : 4; 7677 : 4; Balgooy, M.M.J. van 1887 : 5; 5037 : 4; 5223 : 4; Balgooy, M.M.J. van; Wiriadinata, H. 2865 : 4; Bamler, G. 31 : 4; Bamler, M.G. ROS 132 : 4; Banoc, L.M. 3 : 6; 58 : 6; Banying ak Nyudong S 17211 : 4; S 19419 : 4; Barcelona, J.F. 2040 : 4; Barcelona, J.F.; Busemeyer, D.T. 717 : 4; Barcelona, J.F.; Busemeyer, D.T.; Ippoli, A. 575 : 4; 622 : 4; Bartlett, H.H. 15759 : 4; Beaman, J.H. 6957 : 4; 8027 : 4; 9605 : 4; 9921 : 4; 10332 : 4; Beusekom, C.F. van; Beusekom, R.J. van 1542 : 3; Beusekom, C.F. van; Charoenphol, C. 1690 : 3; Beusekom, C.F. van; et al. 4517 : 3; 4526 a: 3; 4813 : 6; Beusekom, C.F. van; Phengkhlai, C. 2431 : 8; Bor, S. 767 : 8; Borssum Waalkes, J. van 1290 : 4; 2824 : 4; Braithwaite, A.F. 4216 : 9; R.S.N.H. 2370 : 5; RSNH 2087 : 4; RSNH 2110 : 4; RSNH 2436 : 4; RSS 4045 : 9; RSS 4174 : 4; RSS 4471 : 9; RSS 4676 : 5; Brass, L.J. 2916 : 9; 3032 : 4; 3341 : 4; 3893 : 4; 7115 : 4; 11266 : 5; 11870 : 4; 12109 : 5; 12158 : 5; 12841 : 9; 12842 : 4; 13002 : 4; 13214 : 9; 13215 : 4; 13323 : 4; 23016 : 4; 23174 : 5; 24907 : 4; 25803 : 4; 25805 : 4; 26074 : 4; 26087 : 4; 29706 : 5; 31944 : 4; 32064 : 9; Britton, B.B. 326 : 4; Brooke, W.M.A. 8590 : 4; Brooks, C.J. 115 : 4; 263 s: 4; Brown, E.D.W. 980 A: 5; 980 B: 5; 980 C: 5; 980 D: 5; 980 E: 5; Brownlie, G. 1717 : 4; Bünnemeijer, H.A.B. 675 : 4; 3875 : 4; 4127 : 4; 4238 : 4; 5494 : 4; 8969 : 4; 11481 : 3; 12161 : 3; Burkill, I.H. 12904 : 4; SFN 8497 : 4; Buwalda, P. 3640 : 4; 5171 : 4; 6622 : 4; 8063 : 4.
Caerlan; Sageed; Romero PPI 13079 : 4; Carr, C.E. 13428 : 4; 13552 : 5; 13981 : 9; 14419 : 5; 14831 : 9; Carrick, J. JC 102 : 4; Chai, P. S 37561 : 4; S 39456 : 4; Chao Yu Zhang 21441 : 8; Charoenphol, C. 4241 : 3; Cheesman, L.E. 128 : 4; 1199 : 9; Chen Nian-qu 40930 : 2; Cheng Shu-Zhi; Li Bosheng 1015 : 8; 5250 : 4 Chew, W.L. 942 : 4; 1273 : 4; Chew, W.L.; Corner, E.J.H.; Stainton, A. 298 : 4; 1461 : 4; Chin, S.W. 28 : 4; Ching, R.C. 25432 : 8; Christophersen, E. 15 : 4; 125 : 4; 520 : 4; 826 : 5; 866 : 4; 2138 : 5; 3095 : 5; Christophersen, E.; Hume, E.P. 2305 : 4; Chun, N. 40930 : 2; Cicuzza, D. 178 : 4; Clarke, C.B. 42231 A: 6; Clemens, J. 421 : 4; 4534 : 9; 32552 : 5; 32871 : 5; Clemens, J.; Clemens, M.S. 27210 : 4; 28337 : 4; 28763 : 4; 30705 : 4; 30902 : 4; Clemens, M.S. 9114 : 5; 11053 : 4; 16494 : 4; 41009 : 5; Clunie, N.M.U. LAE 63276 : 9; LAE 63380 : 9; LAE 63399 : 4; Co, L. 3197 : 4; Coert, J.H. 644 : 4; 1497 : 4; Collecteur de Darjeeling 109 : 8; Coode, M.J.E. 6498 : 4; NGF 32846 : 4; Coode, M.J.E.; et al. 7548 : 4; Copeland, E.B. 137 : 4; 1381 : 4; 1474 : 4; 1766 : 4; 1804 : 2; 2016 : 4; ppe 56 : 4; Cox, P.A. 224 : 4; 345 : 4; 838 : 4; Craven, L.A. 1123 : 5; 1269 : 5; Craven, L.A.; Schodde, R. 133 : 4; 287 : 9; Croft, J.R. 34 : 4; 66 : 4; 224 : 9; 817 : 4; 821 : 4; 1223 : 4; 1897 : 9; 1987 : 9; Croft 11 : 5; Croft 875 : 5; Croft 1508 : 5; LAE 65781 : 5; LAE 65822 : 5; LAE 68337 : 5; LAE 68911 : 4; LAE 71012 : 4; LAE 71112 : 4; Croft, J.R.; Katik, P. NGF 14933 : 4; Croft, J.R.; Lelean, Y. LAE 65642 : 4; Croxall, J.P. 6016 : 5; Cuming, H. 60 : 2; 94 : 4; Curry, P. 1644 : 4; Curtis, C. 160 : 4.
Damas, K. LAE 58855 : 4; Danser, B.H. 5971 : 4; Darbyshire, P.J. 226 : 7; 325 : 9; Darnaedi, D. 1554 : 5; 2362 : 4; 2725 : 3; Davidse, G.; Sumithraarachchi, D.B. 7965 : 3; Davis, A.P. 464 : 4; 814 : 5; 814 : 5; Degener, O. 14279 : 4; Docters van Leeuwen, W.M. 10942 : 4; Dodd, J. E 10 : 9; Dransfield, S. 976 : 4.
Ecology Highland Group 14053 : 8; Edaño, G.E. 26 : 4; 71 : 4; 303 : 4; 637 : 4; 647 : 4; 796 : 4; 806 : 4; 1540 : 4; 2596 : 4; 5922 : 4; 6260 : 4; 8031 : 4; pnh 17267 : 4; Edwards, P.J. 724 : 1; 1988 : 4; 2141 : 4; Elbert, J. 1650 : 3; Elmer, A.D.E. 7940 : 4; 9069 : 4; 9959 : 4; 10908 : 4; 11451 : 5; 14140 : 4; 14149 : 4; 17694 : 4; 17964 : 4; Endert, F.H. 3030 : 4; 4244 : 4; Evans, J.H.N.; Gordon, W. 998 : 2; Eyma, P.J. 1664 : 5; 4484 : 9; 4726 : 4.
Fallen, M.E. 437 : 5; Faurie, U. 611 : 8; Florence, J. 3634 : 5; 4336 : 5; 6780 : 5; 7498 : 5; 9623 : 5; 9770 : 5; Forbes, H.O. 662 : 3; 884 : 3; 3482 : 2; Foreman, D.B. LAE 59150 : 9; NGF 45759 : 4; forest production group 84422 : 2; Forrest, G. 11799 : 8; 18581 : 8; 24238 : 8; 26691 : 8; Fosberg, F.R. 62670 : 5; Foxworthy, F.W. 72 : 4; 77 : 4; 359 : 4; Fu Guoxun 415 : 8; Fuchs, H.P.; Collenette, S.H. 21671 : 4.
Gaerlan, F.J.M.; Sagcal, E.; Romero PPI 10874 : 4; PPI 13079 : 4; Gagné, B.H. 1113 : 5; 1580 : 5; BHG 2318 : 5; Garber, D.W. 724 : 4; 1046 : 4; Gawi, M. 11 : 5; Gebo, A. 1619 : 4; Geesink, R.; Santisuk, T. 5384 : 2; Ghose, G. 11 : 3; Gideon, O. LAE 57504 : 4; Gideon, O.; Silu, J. LAE 76918 : 4; Gillespie, J.W. 2748 : 4; 3249 : 4; 3815 : 4; 4110 : 5; Gillett, G.W. 2174 : 5; Gong Wu Su 479 : 8; Graeffe 1078 : 4; Grant, M.L. 3704 : 5; 4095 : 5; Gravendeel, B. 593 : 4; Grey; Wilson, E.H.; Phillips 289 : 3; Gurung, V.L. 55 : 2; Gwynne Vaughan, D. 431 : 4.
Haas, J.H. de 2622 : 8; Hallier, H. 418 : 4; 1699 : 4; 3297 : 4; Hansen, B.; Smitinand, T. 11846 : 2; Hansen, C. 1128 : 4; Hardeveld, C. van 318 : 3; Hartley, T.G. 11417 : 4; Harvey, H.D. 1668 : 4; Hassan Flora Project 579 : 3; 692 : 3; 887 : 3; Henderson, M.R. 11197 : 4; 17879 : 4; Hennipman, E. 3092 : 3; 3334 : 6; 3334 A: 2; 3334 B: 6; 3413 : 8; 3568 : 3; 3655 : 3; 5370 : 5; 5430 : 5; 5518 : 4; 6158 : 4; Henry, A. 9484C : 2; 9484 : 8; Hirano, M.; Hotta, M. 1522 : 4; Hirschland, J.G. 2 : 4; Hochreutiner, B.P.G. 1755 : 4; 3326 : 4; Hodel, D.R. 1373 : 5; Höft, R. 2260 : 5; 2733 : 5; 2953 : 5; 3659 : 5; Holstvoogd, C. 471 : 4; 472 : 3; 847 : 3; 848 : 3; Holttum, R.E. 1668 : 4; 20580 : 4; 31319 : 4; N.G.F 40181 : 5; SFN 10686 : 4; SFN 11403 : 4; SFN 14852 : 2; SFN 25723 : 4; SFN 39209 : 3; Hoover, W.S. 434 : 9; Hortus, Bogor 57 : 4; 177 : 4; Hosokawa, Takahide 8677 : 4; Hotta, M. 14797 : 1; Hou, D. 213 : 4; Hovenkamp, P.H. 527 : 4; 05 05 : 4; Huang, T.C.; Kao, M.T. 1730 : 8; Hume, H.L. 7174 : 4.
Investigation team in western Yunnan 11416 : 8; Iwatsuki, K. 484 : 8; 531 : 3; B 1870 : 4; B 2495 : 4; B 2505 : 4; C 921 : 9; P 597 : 3; Iwatsuki, K.; Fukuoka, N. T 3424 : 6.
Jaag, O. 1110 a: 3; 1170 : 3; 1960 : 4; Jacobs, M. 7948 : 4; Jacobson, E. 2549 : 4; Jermy, A.C. 3488 : 5; 13677 : 4; 14420 : 1; Jiang Xinglin 35239 : 8; John, H.; St 17025 : 5; Johns, R.J. 7369 : 4; 7986 : 5; 9023 : 5; 9091 : 5; 10303 : 4; 10329 : 5; Jones, W.B. 1798 : 5.
Kadim bin Tassim K 491 : 2; Kajewski, S.F. 874 : 4; 2691 : 4; Kalkman, C. 4005 : 4; 4379 : 5; 4469 : 5; Kathmandu Expedition KE 1067 : 8; Katik, P. LAE 56318 : 4; Kato, M. 822 : 9; B 3674 : 4; B 6042 : 4; B 7667 : 4; B 7901 : 4; B 9511 : 1; B 9569 : 1; B 9813 : 1; B 10920 : 1; B 11131 : 4; C 1278 : 9; C 1365 : 4; C 1729 : 4; C 1945 : 4; C 2907 : 4; C 4121 : 4; C 5058 : 4; C 7412 : 9; C 7480 : 5; C 11656 : 4; C 12846 : 5; C 12965 : 4; C 13351 : 4; C 14279 : 4; C 14361 : 5; C 14446 : 4; Kato, M.; Setoguchi, H.; Darnaedi, D. 1160 : 4; Kato, M.; Wiriadinata, H. B 4749 : 4; B 4924 : 4; B 5283 : 4; B 5931 : 4; Kaudern, W. 86 : 4; Keenan 1386 : 6; Kerr, A.F.G. 9363 : 6; 9645 : 3; Kessler, P.J.A. 2857 : 4; Keysser, C. II 74 : 5; Kinbag, F. 013 : 5; King’s collector 2424 : 4; Kog, P. 004 : 5; Koorders, S.H. 14942 B: 4; 19832 b: 3; 36678 B: 4; Koster, C. BW 13848 : 4; Kostermans, A.J.G.H. 2280 : 4; 5549 : 4; 6513 : 4; Kostermans, A.J.G.H.; Soegeng, W. 509 : 4; Kostermans, A.J.G.H.; Wirawan, N. 747 : 3; Kuhl, H.; Hasselt, J.C. van 7 : 4; Kurz, S. 203 : 3.
Lace, J.H. 4227 : 6; LAE 65715 : 4; Lai, J.; Enjah S 64162 : 4; Lam, H.J. 537 : 4; 1449 : 4; 1856 : 5; 1943 : 5; 6897 : 4; Lang Kai-Yong; Zhang Yongtian 3360 : 8; Larsen, K. 44 : 3; 964 : 3; 2689 : 6; 8936 : 6; 10678 : 3; 31861 : 6; Larsen, K.; Larsen, S.S. 30713 : 3; Lau S.K. 1490 : 2; 3108 : 2; 27368 : 2; Lavarack, P. NGF 31446 : 5; Lavarack, P.; Ridsdale, C.E. NGF 31444 : 9; LeBronnec, G. 809 : 5; Ledermann 7652 : 7; Leeuwenberg, A.J.M. 3 : 4; Li Bosheng 14023 : 8; Li Hua Hou 1009 : 2; Li Yang Yi; Li Bosheng 13537 : 8; Liu Bingrong 5064 : 2; 5065 : 2; Lorence, D.H. 6094 : 5; Lörzing, J.A. 728 : 3; 5202 : 4; 6890 : 4; 14050 : 4; Luang Winit-Wanadorn 47 : 6; Luang, Winit 1003 : 3; Ludlow, F. 6496 : 8; 6685 : 8; Ludlow, F.; Sherriff, G.; Hicks, J.H. 17008 : 8; 17416 : 8; 17499 : 8; 17536 : 8; 17540 : 8; 19563 : 8; 20909 : 8; Luerssen 3766 : 4; 9902 : 4.
M. K. Li 450 : 2; MacDaniels, L.H. 1531 : 5; Main 261 : 4; Mamit, J.D. S 34391 : 4; Mangen, J.M. 2227 : 4; Manickam, V.S.; Matthew, K.M. RHT 34242 : 3; Manner; Street 346 : 4; 592 : 4; Manseima, J. 011 : 5; Martellino, A.; Edaño, G.E. BS 35643 : 4; Maxwell, J.F. 85-194 : 4; 74 777 : 3; 74 907 : 6; 76 571 : 3; 78 167 : 4; 85 495 : 4; 87 227 : 2; 87 641 : 6; 89 858 : 6; 91 495 : 6; 92 716 : 6; 93 774 : 3; 93 1359 : 6; 94 1030 : 8; 94 1303 : 6; 95 919 : 6; 96 1265 : 3; 97 720 : 6; 97 1082 : 6; 98 636 : 6; McClure, F.A. 20061 : 2; McGregor, R.C. BS 10316 : 4; McKee, H.S. 3054 : 5; Meebold 7512 : 6; Mei Fung 4879 : 8; Meijer, W. 69 : 3; 905 : 4; 9473 : 4; 10341 : 3; Mendoza, D.R.; Convocar, P. 712 : 4; 748 : 4; Merrill, E.D. 624 : 4; 3238 : 4; 5937 : 4; 6682 : 2; Sp. Blan 490 : 4; Middleton, D.J.; Meng Monyrak 617 : 4; Millar N.G.F. 15745 : 5; Mogea, J.P.: Wilde, W.J.J.O de 3715 : 4; Mohd Haniff SFN 8019 : 4; Mohd Nur SFN 34358 : 4; Mohd Shah MS 656 : 4; MS 2803 : 4; Molesworth Allen, B. 4574 : 2; Moulton, J.C. SFN 6764 : 4; Moysey, L. 31884 : 4; Mumford, E.P. 472 : 5; Murata, G. J 890 : 4; T 16057 : 8.
Nakaike, K. 558 : 5; native collector 2222 : 4; SFN 4179 : 2; Neervoort, A.M. 253 : 4; Nitta, A. 15091 : 3; Normal School Students 12691 : 4; Norris, W. 856 : 4.
Oliver, R.L. 3109 : 5; Ooststroom, S.J. van 14090 : 3.
Pacific Entomological Survey 474 : 5; Ex 472 : 5; Palmer, W. 512 : 4; Palmer, W.; Bryant, O. 254 : 3; Parks, H.E. 20759 : 4; 70904 : 4; Parris, B.S. 7913 : 5; 10550 : 4; 11252 : 4; 11331 : 4; Parris, B.S.; Croxall, J.P. 5968 : 4; Parris, B.S.; Edwards, P.J. 10479 : 2; Parry Hance 1998 : 2; Pearce, K.G.; Serukit, D. S 95467 : 1; Perlman, S. 10118 : 5; Petrmitr, O. 164 : 6; 397 : 6; Phengkhlai, C. 283 : 2; 305 : 6; Phoon, S.N. FRI 53265 : 4; Pierre, L. 5713 : 4; Piggott, A.G. 2187 : 4; Pleyte, D.R. 343 : 4; Poilane, E. 4219 : 3; 9540 : 6; 12673 : 8; 15805 : 2; 23031 : 4; Polak, A.M. 931 : 4; Polak, E. 2048 : 4; Pooma, R. 4830 : 6; Pooma, R.; et al. 4373 : 4; Posthumus, O. 2429 : 4; 4058 : 4; Price, M.G. 1157 : 4; 2691 : 4; Price, M.G.; Hernaez, B.F. 701 : 4; Pulle, A.A. 113 : 4; 484 : 4; 596 : 4; 722 : 5; 1086 : 5; Pullen, R. 7738 : 4; 8069 : 4.
Qi Xinping Q 77 : 8; Qinghai-Tibet Team 4427 : 8; 4920 : 8; 8673 : 8; Qiu Yun 52907 : 8; Quayle, E.H. 61 : 5; 1145 : 5; 1258 : 5.
Raap, H. 218 : 4; 227 : 4; Rachmat 496 : 4; Ramos, M. 14881 : 4; Raynal, J. 17073 : 4; Raynal, J.; Schmid, R. RSNH 16123 : 4; Reinecke, F. 108 : 4; Reinecke, F.? 94 : 4; Reynoso; Garcia; Sagcal, E. PPI 14300 : 4; Ridley, H.N. 571 : 2; 5171 : 2; 5172 : 2; 7832 : 4; Riswan, S.; Afriastini, J.J. J 075 : 4; Robinson, H.C. 6024 : 4; Rock, J. 2026 : 6; Römer, L.S.A.M. von 770 : 4; Royen, P. van 3644 : 4; 3842 : 4; 3870 : 5; 7373 : 5; Royen, P. van; Sleumer, H.O. 5959 : 9; 6344 : 4; 7485 : 4; 7651 : 4; Rutten 1567 : 4; 2047 : 9; 2198 : 9; 2244 : 4.
S. Y. Dong 136 : 3; Sachet, M.-.H. 1021 : 5; Saigol, P. SAN 93067 : 4; Saldanha 17990 : 3; Sands, M.J.S. 6399 : 4; Sauveur; Sinke 2516 : 4; Schiffner, V. P 209 : 4; Schlechter, R. 16689 : 9; 18572 : 4; 19626 : 4; Schmutz, E. 59 : 3; 636 : 3; 2282 : 3; 6086 : 2; F 24 : 2; Schnell, R. 10176 : 3; Schwartz, A. 2448 : 4; 2795 : 3; 2822 : 4; Scortechini, B. 394 : 4; Seimund, E. 236 : 4; Setchell, W.A. 398 : 4; Shimizu, T. M 13244 : 4; T 8959 : 2; T 11457 : 3; T 22791 : 2; Shui Yumin 3730 : 8; Sidiyasa, K.; Arifin, Z. 1586 : 4; Siew Wei Hoe 125 : 4; Simpson, D.A. 2306 : 4; Sinclair, J. SFN 38688 : 4; Sino-Japanese expedition 9391 : 8; T 273 : 8; Sino-Soviet joint mission in Yunnan 2439 : 8; 3798 : 8; 4494 : 8; Sledge, W.A. 1106 : 3; 1790 : 5; Sleumer, H.O.; Vink, W. BW 14084 : 4; BW 14245 : 4; Smith, A.C. 283 : 4; 1870 : 4; 1975 : 4; 4695 : 4; 4811 : 4; 5162 : 4; 5163 : 5; 5423 : 4; 6104 : 4; 8369 : 4; 8502 : 4; 8678 : 4; Smith, E. 894 : 4; 1073 : 3; 1074 : 3; 1281 : 6; 1283 : 6; Smith, L.S. 4602 : 3; Soegeng, W. 433 : 4; Soejarto, J.J.; Fernando, O.; Sagcal, E. 8874 : 4; Soepadmo, E.; Mahmud 1091 : 4; Steenis, C.G.G.J. van 3688 : 4; 18293 : 3; 18504 : 2; 18505 : 4; 20870 : 8; Stevens, P.F. LAE 50373 : 4; LAE 58317 : 9; LAE 58759 : 9; Stevens, P.F.; Lelean, Y. LAE 58693 : 4; Stewart, R.R. 15949 : 8; Stone, B.C.; Price, M.G. 12190 : 4; Stone, B.C.; Reynoso; Sagcal, E. PPI 447 : 4; Streimann, H. N.G.F. 35860 : 5; NGF 28939 : 4; Streimann, H.; Kairo, A. NGF 39009 : 4; NGF 44042 : 9; Stresemann, E. 9 : 4; 12 : 9; 365 : 9; 405 : 9; 405 a: 5; Strugnell, E.J. 14611 : 4; Sulit, M.D. 1211 : 4; 5376 : 4; Sun Hong Fan Herb 9434 : 3; Surbeck, H. 152 : 4; 600 : 3; 1036 : 4; 1093 : 4; Survey team who collected herbs in Tibet 1102 : 8; 1102 : 8; 1139 : 8; Swart, J.W. 2448 : 4.
Tagawa, M. 542 : 8; T 634 : 3; T 646 : 2; T 1524 : 3; T 9311 : 3; Takeuchi, W. 5340 : 9; 5390 : 9; 6229 : 4; 6317 : 9; 6318 : 4; 8920 : 4; 9234 : 4; 10703 : 5; 10758 : 4; 10775 : 4; 10814 : 9; 11714 : 4; 12970 : 4; Takeuchi, W.; Towati, A.; Ama, D. 19917 : 5; Teck, L.S. S 68652 : 4; Thwaites, G.H.K. CP 1378 : 3; Ting, K.C. 796 : 2; Topping, D.L. 450 : 4; 850 : 4; 1338 : 4; 1856 : 4; Tsiang, Y. 4731 : 2; Tsoong, K.K. 1284 : 2; Turnau, E.A. 846 : 4.
Ueda, K.; Darnaedi, D. B 8941 : 4; University of San Carlos 79 : 4; 120 : 4; Unknown 46-2 : 4; 56 : 8; 60 : 2; 4699 : 8; 13924 : 4; 15513 : 4; Utteridge, T.M.A. 381 : 5.
Vaupel, F. 337 : 4; Vegetation team Qinghai-Tibet group 2503 : 8; Veldkamp, J.F. 8446 : 4; Versteeg, G.M. 1335 : 4; Versteegh, C. BW 10299 : 4; BW 10419 : 4; BW 10420 : 5; BW 10424 : 9; BW 12561 : 4; Vink, W. 17596 : 5; Vogel, E.F. de 1939 : 4; Vogel, E.F. de; Vermeulen, J.J. 7310 : 4; 7561 : 4; Voogd, C.N.A. de 128 : 4; 2498 : 3; Vriese, W.H. de 6 : 3; 25 : 3; 26 : 4; 28 : 3; 35 : 4; 66 : 4; 342 : 3.
Wagner, W.L. 6225 : 5; Waitz, F.A.C. 27 : 4; Walker, T.G. 8042 : 9; 8265 : 9; T 7541 : 5; Wallich, N. 373 : 8; Wang, C.W. 39287 : 3; 39445 : 8; 74763 : 6; 78515 : 3; 87504 : 2; Wang, W.T. 10419 : 6; Wang, Z.R. C 621 : 2; Watthana, S. QBG 21791 : 3; Webster, G.L.; Hildreth, R. 14181 : 4; Werner, E. ROS 28 : 9; Western China Academy of Sciences 4699 : 8; Whistler, A. 3549 : 4; Whitford, H.N. 999 : 4; 2248 : 4; Whitmore, T.C. BSIP 997 : 9; BSIP 2133 : 9; Whitmore, T.C.; Womersley, J.S. BSIP 688 : 9; Widjaja, E.A. 4207 : 4; Wilde, A.G. de; Vervoort, W. 530 : 4; Wilde, W.J.J.O. de; Wilde-Duyfjes, B.E.E. de 12942 : 4; 13787 : 4; 15022 : 4; Williams, K. 1981 : 4; Wilson, E.H. 5246 : 8; Winckel, W.F. 967 B: 4; 1359 : 4; 1656 B: 4; Winkler, H. 467 : 4; 1078 : 4; Wiriadinata, H. 144 : 4; Womersley, J.S.; Millar, A. NGF 7775 : 4; NGF 8377 : 4; Wong, K.M. 459 : 4; 1122 : 4; 1444 : 4; FRI 32229 : 4;; Wood, K.R. 4442 : 5; 4583 : 5; 6387 : 5; Worthington, R.D. 12919 : 4; Wray, L. 585 : 4; 1601 : 4; 3729 : 4; 5357 : 4; Wuzhishan Fern Survey 217 : 3; Wyatt-Smith, J. KEP 78837 : 4.
Xinggong Xia Xia 5419 : 8; 5520 : 8; Xu 4 : 8.
Y. Tsiang 11574 : 8; Yahud; Mahmud; S 88396 : 1; Yates, H.S. 2899 : 4; Yii Puan Ching SAR 48495 : 4; Yoshida, S. 1552 : 4; Yan Yue-Hong 3101 : 2; 12943 : 2; Yu, T.T. 16648 : 8; 17303 : 8; 17305 : 8.
Zhang Xian-Chun 106 : 2; 2355 : 3; Zhang Xian-Chun; Chen Zhen Chuan 3781 : 2; Zhang Xian-Chun; Dong Shiyong 1404 : 6; 1423 : 6; Zhongshan University 23749 : 8; Zhu Taiping 189 : 6; 476 : 6; Zhu Weiming 2705 : 8; Zollinger, H. 507 : 3; 1306 : 4; 1306 ?: 4; 1306 a: 3; 1306 B: 4; 1993 : 3.