





(C) 2011 Robert B. Faden. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Polyspatha oligospatha Faden, the third species in a small African endemic genus of Commelinaceae, is described. It is widespread but has been overlooked because of its small stature and resemblance to small plants of Polyspatha paniculata. It differs from both Polyspatha paniculata and Polyspatha hirsuta, the two other species, by its leaf pubescence, fewer, more widely spaced and usually patent spathes, deeply ridged seeds with numerous knobby, transversely interrupted ridges, and morning anthesis. It occurs throughout the Congolian forests from Cameroon to Uganda, but it is also disjunct in Ivory Coast, across the Dahomey gap.
Cameroon, Dahomey gap, seed morphology, Uganda, Ivory Coast, morning anthesis, disjunct distribution
The African endemic genus Polyspatha (Commelinaceae) has generally been considered to consist of two species (
When I first began working on the Commelinaceae of Cameroon and examined the Polyspatha specimens at the Herbier National du Cameroun (YA) in Yaoundé, Cameroon in 1986, it appeared that there were three distinct species of Polyspatha, not the usually accepted two. I came to the same conclusion from studies at the Muséum National d’Histoire Naturelle, Paris (P) the following year. Later studies at the Royal Botanic Gardens Kew (K), in connection with work on the Flora of Tropical East Africa, confirmed that the same three species also occurred in Uganda, some 2000 km away. Studies of collections from other herbaria, particularly the National Botanic Garden of Belgium, Brussels (BR), confirmed that the three species also were present in the intervening territory. It was particularly significant that this species was also found to occur in Ivory Coast, almost 1900 km from the nearest locality in Cameroon.
MethodsMore than 540 herbarium specimens of Polyspatha
were studied from the following institutions: all the collections from
BM, BR, EA, K, NY, MO, US, WAG and YA, and some of the
collections from P. Living material of Polyspatha oligospatha (Poulsen 1275, originally from Uganda, and Faden et al. 86/2,
originally from Cameroon) was cultivated at the Smithsonian
Institution Botany Research Greenhouse, in Suitland, Maryland, USA
and provided data on flowering times, root tips for chromosome counts,
seed collections and liquid preserved flowers. A mitotic chromosome
count was obtained using the techniques of
Polyspatha oligospatha Faden sp. nov. a Polyspatha paniculata foliis trichomatibus uniseriatis in pagina superiore instructis, spathis plerumque minus numerosis differt; a Polyspatha hirsuta pubesentia foliorum plerumque multo sparsa trichomatibus uniseriatis in pagina inferiore semper carentibus, spathis minus congestis minus numerosis minus pubescentibus et plerumque patentibus differt; et ab ambo ordinatione testae et anthesi mane differt. Type: Uganda, Zintengese [=Zintengeze], (Mabira), September 1922, R. A. Dummer 5531 (US!, holo, K!, iso).
Stoloniferous perennial with erect shoots
5–20(-30) cm tall; internodes puberulous with hook-hairs, very rarely
also with short, uniseriate hairs. Leaves usually subclustered
terminally on the flowering shoot, sheaths 0.8–1.5 cm long, puberulous
with hook-hairs (

Polyspatha oligospatha Faden, sp. nov. 1. Habit. 2. Spathe with open flower, side view. 3. Flower, front view. 4. Flower, side view. 5. Stamen and staminode filaments, showing basal fusion. 6. Dehisced capsule. 7. Seed, dorsal view. 8. Seed, ventral. All from Poulsen 1275 (originally from Uganda; cultivated at the Smithsonian Institution). Illustration by A. R. Tangerini.
Ivory Coast, Cameroon, Republic of Congo (Congo-Brazzaville), Democratic Republic of Congo (Congo-Kinshasa), Sudan, Uganda (Figure 2).
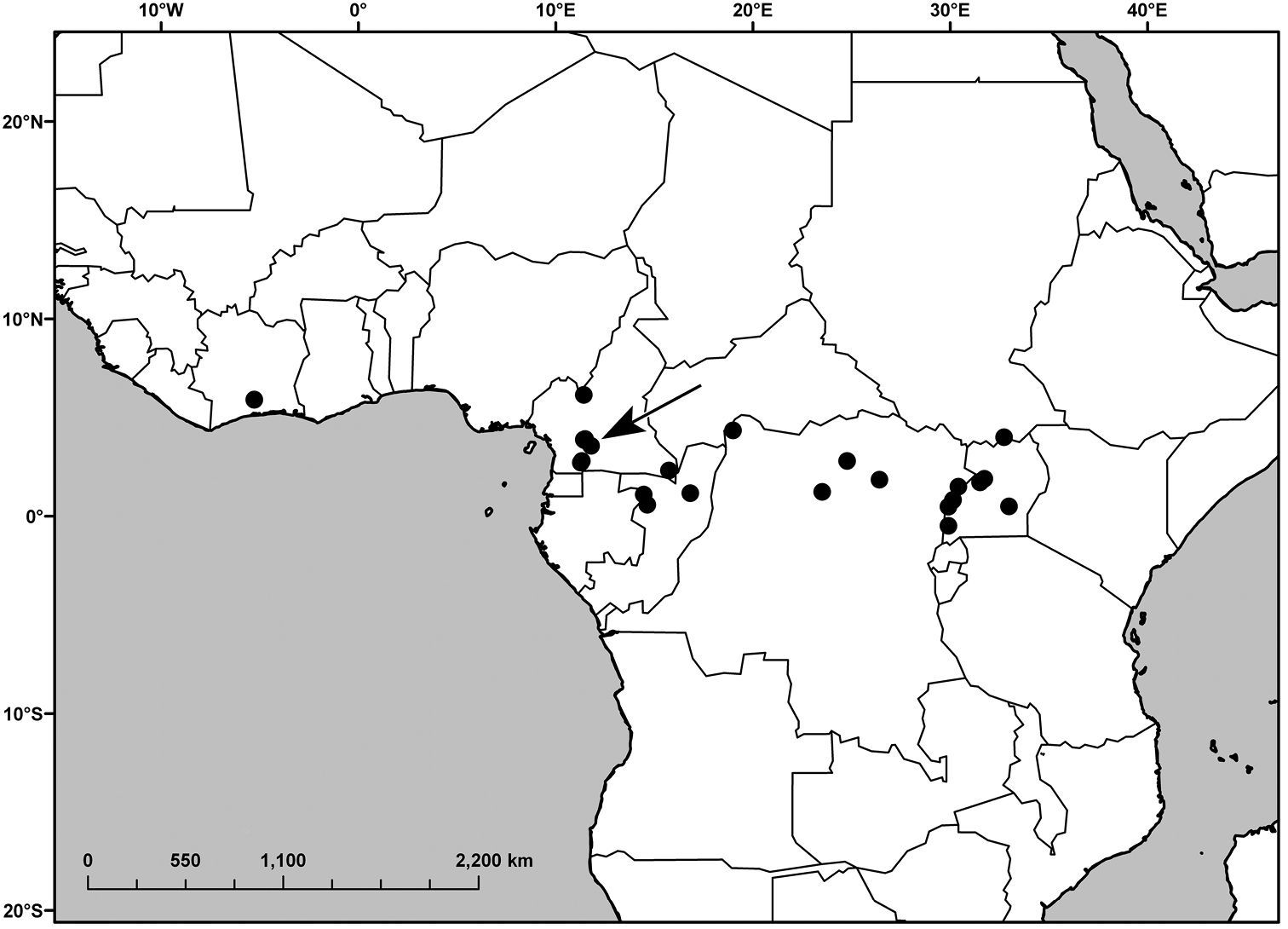
Distribution of Polyspatha oligospatha Faden, sp. nov. The arrow indicates the location of the collection Letouzey 11465, which is atypcial for the species (see text).
Understory in evergreen and semi-deciduous forests, forest relicts and other moist, shaded habitats in savanna, shaded cultivation, especially of cacao; ca. (15-)600–1220 m.
2n = 28 (from Faden et al. 86/2 from Cameroon, cultivated at the Smithsonian Institution [Faden unpublished]).
A total of 24 flowering collections of Polyspatha oligospatha have been seen, with specimens collected from all months except April and May. Anthesis occurs in the morning, based on the following records. Poulsen 1275 from Uganda, cultivated at the Smithsonian Institution Botany Research Greenhouse in 1997, had flowers open by 9:30 a.m. and fading at noon. Two collections from Cameroon, Hall & Kahn 075/93 and Keating 90–13 (both US), record this species as flowering in the morning and fading about noon.
Polyspatha oligospatha is easily recognizable but also readily overlooked, judging by its wide distribution and modest number of collections. It is distinctive because of its small stature, leaves with long, uniseriate hairs only on the adaxial surface, small inflorescences with small, well-spaced, often patent (i.e. not becoming deflexed) spathes whose surface is composed entirely or partly of lustrous, brown, bead-like cells, and seeds with numerous, radiating, knobby ridges that are transversely interrupted (Figure 1: 7, 8; Plate 1: 3, 4).
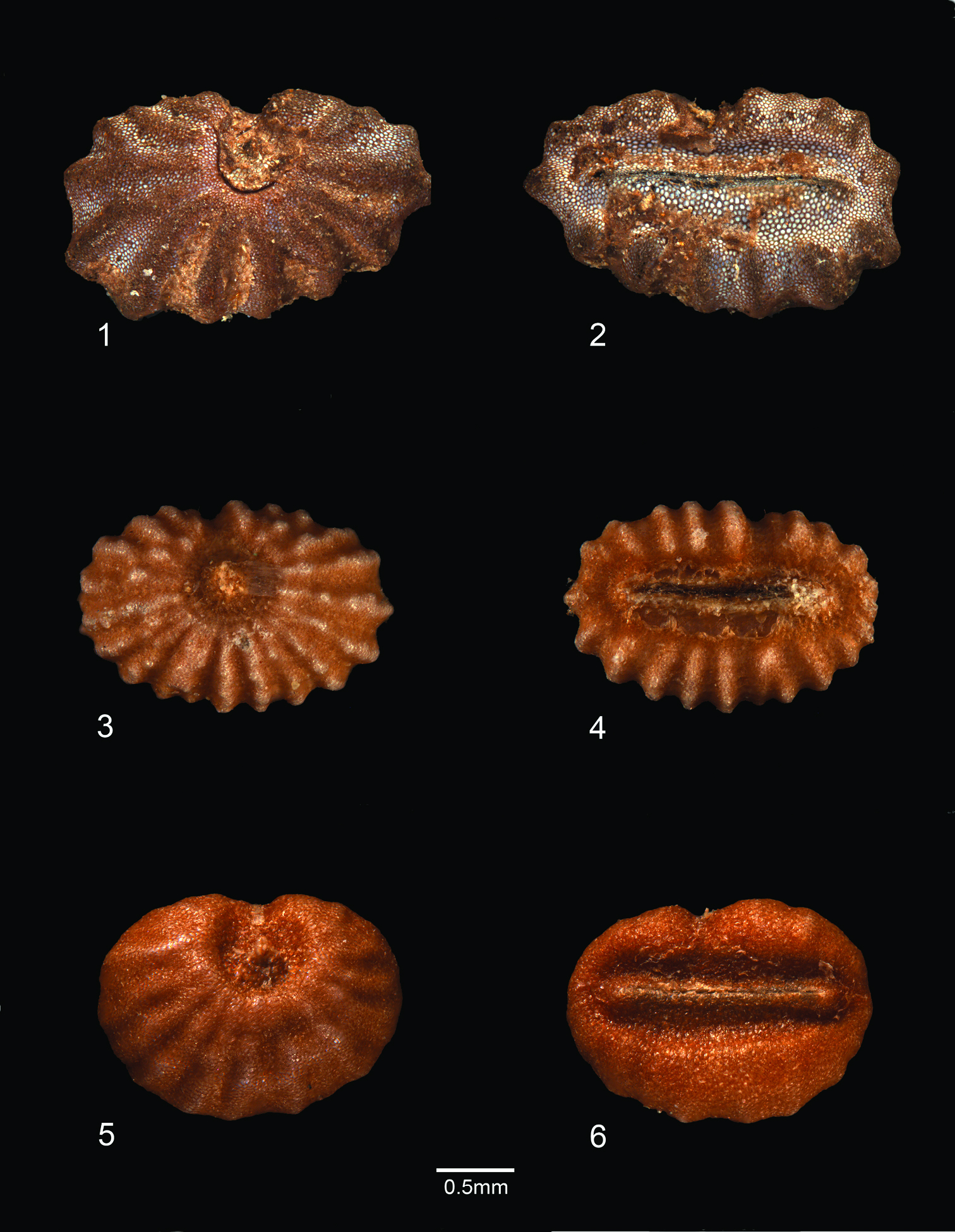
Seeds of Polyspatha species 1–2 Polyspatha paniculata Benth. 1 Seed, dorsal view 2 Seed, ventral view. (From Faden et al. 74/35 from Ghana, cultivated at Smithsonian Institution) 3–4 Polyspatha oligospatha Faden, sp. nov. 3 Seed, dorsal view. 4. Seed, ventral view. (From de Wilde & de Wilde-Duyfjes 2647 [BR] from Cameroon) 5–6 Polyspatha hirsuta Mildbr. 5 Seed, dorsal view 6 Seed, ventral view. (From Kahn 92-1 [US] from Cameroon).
Adamoua Region: Near Kongi (10 km NW of Kimi-Bankim on Foumban - Banyo route), 27 June 1967, Letouzey 8742 (K, P, WAG) [two additional sheets of this collection at P are Polyspatha paniculata]. Center Region: N’Kolbisson, ca. 8 km W of Yaoundé, 2 Dec. 1963, de Wilde & de Wilde-Duyfjes 1369 (P, WAG); same locality, 7 June 1964, de Wilde & de Wilde-Duyfjes 2647 (BR, K, WAG, YA); Nkolbisson, 8 km W of Yaoundé, Akouandoué Hill S of the town, 23 Jan.1986, Faden, Satabié & Mpom 86/2 (K, P, US, YA); Nkolbisson, 8 km W of Yaoundé, 3 km NW of the town, 24 Jan. 1986, Faden, Satabié & Mpom 86/22 (US); Mt. Febe, Yaoundé, 4 July1987, Manning 2166 (K, MO, WAG); Colline au N de Nkolbison (8 km W Yaoundé), 6 Feb.1963, Raynal & Raynal 9445 (P, YA). East Region: Boumba-Ngoko Dept., Moloundou Arrondissement, Lobeke swamp/savanna, adjacent to camp (site of an old sport hunting camp), 6 Feb.1993, Hall & Kahn 075/93 (K, P, US, YA, WAG). South Region: Ntem Departement, Ako’okas, 40 km SE of Ebolowa (by road), 30 June 1993, Keating 90–13 (US); same locality, 4 Aug. 1993, Keating 90–13 bis (K, P, US, YA); Rocheur de Mokomessi, 20 km NNW of Zoetélé, 35 km NNW of Sangmelima, 9 July1992, Letouzey 11465 (BR, K, P, YA); Ebolowa - Si I (W d’Ebolowa), basses pentes d’une colline dominant la ville, 9 Mar. 1963, Raynal & Raynal 10340 (P, YA).
Bassin de l’Alimo-Likoula, Réserve de Chasse de M’Boko, salime au bord de la Lekoli près du campement, 10 Aug. 1961, Descoings 9059 (P); Grand escarpement d’Odzala, au Nord de la cascade dorée, 27 Nov. 1996, Lejoly 96/961 (BR); Inter Bonga et Wesso (Sanga), Aug. 1899, Schlechter 12714 (BR) [mixture with Polyspatha paniculata; K sheet of this number is pure Polyspatha paniculata].
Forestier Central (VI):en aval de Barumbu, 27 Oct. 1913, Bequaert 971 (BR); Panga, 19 Dec. 1913, Bequaert 1561 (BR); Uelé, Route Buta – Buna, 15 Oct. 1905, Seret 70 (BR) [2 sheets at BR, each with one plant of Polyspatha oligospatha (renumbered Seret 70A) and one of Polyspatha paniculata (renumbered Seret 70B); mapped as “Buta”)]. Ubangi-Uele (VII): Zongo (Ubangi), Nov. 1930, Lebrun 1729 (BR). Lacs Edouard et Kivu (IX): Vallée de la Semliki, a l’est de Beni, July 1929, Humbert 8796 (BR, P).
Divo Forest, Oct. 1959, Aké Assi 5707 (K); North Divo Forest Reserve, 12 Oct. 1959, Fosberg 40559 (K, US).
Equatoria Province, Torit District: Tallanga Forest, 28 Dec. 1949, Jackson 1005 (BM).
Bunyoro (Masindi) District: Pabidi Forest, 14 Sep. 1995, Poulsen et al. 958 (C, US); Budongo Forest Reserve, Kanyo-Pabidi Block, 4 Feb. 1996, Poulsen et al. 1180 (C); Budongo Forest Reserve, between the Royal Mile and the Nature Reserve, close to the border of the Nature Reserve, at the campsite at the Nyabisabo River (gap), 4 Dec.1996, Poulsen et al. 1245 (US) & 1246 (K, MO); Budongo Forest, 17 July 1969, Stewart in EAH14170 (EA, K). Kigezi District: Just outside Queen Elizabeth Nat. Park, South Maramagambo Central Forest Reserve, 6.4–11.2 km up Kaizi-Bitereko road which is off the Katunguru-Ishsha road (Congo road), 18 Sep.1969, Faden et al. 69/1113 (BR, EA); South Maramagambo Central Forest Reserve, just outside Queen Elizabeth Nat. Park, 8 June 1969, Lock 69/155 (EA); Maramagambo Forest, 18 Sep.1969, Lye et al. 4117 (K). Mengo District: Zintengese [=Zintengeze], (Mabira), Sep. 1922, Dummer 5531 (K, US). Toro District: “Semliki District, ” 31 Oct. 1905, Daws 683 (K); Bwamba County, 1.2 km S of Sempaya, 23 Sep. 1969, Faden, Evans & Lye 69/1256 (EA); Bwamba, Buyayu-Sempayo road, Oct. 1929, Liebenberg 922 & 922A (K) [both are mixtures with Polyspatha paniculata].
| 1a | Lamina lacking long, uniseriate hairs, often scabrous above; seeds ribbed, with (12-)14–18 smooth, uninterrupted ribs | Polyspatha paniculata |
| 1b | Lamina with long, uniseriate hairs, at least on the adaxial surface, never scabrous above; seeds either shallowly ribbed-reticulate or deeply ribbed with (17-)18–23 prominent, knobby ribs that are transversely interrupted | 2 |
| 2a | Long, uniseriate hairs present on the adaxial leaf surface and summits of the leaf sheaths, always lacking from the abaxial leaf surface and usually elsewhere; spathes well spaced, usually patent to slightly deflexed; cells of the spathe surface, at least near the midrib, but often all over, lustrous, brown, and bead-like under a 20× lens; seeds deeply ribbed, the ribs knobby and transversely interrupted | Polyspatha oligospatha |
| 2b | Long, uniseriate hairs usually present on both leaf surfaces, the internodes and sheath surfaces (lacking only in some collections from Nigeria and further west); spathes crowded, becoming deflexed against the inflorescence axis; cells of the spathe surface dull, neither brown nor bead-like under a 20× lens; seeds shallowly ribbed-reticulate, the ribs neither knobby nor transversely interrupted | Polyspatha hirsuta |
Polyspatha oligospatha [etymology: many spathes, few spathes] differs from Polyspatha paniculata by the consistent presence of long, uniseriate hairs on the adaxial leaf surface (vs. none), which is never scabrous (vs. often scabrous) and by the usually fewer, smaller, blunter, more widely spaced and less deflexed spathes. It differs from Polyspatha hirsuta by the usually sparser and shorter uniseriate hairs on the adaxial leaf surface, the absence of such hairs from the abaxial leaf surface and usually the internodes and sheaths (except for the sheath summit), and the inflorescence consisting of fewer, less congested spathes with shorter marginal hairs and with lustrous, brown, bead-like cells on the entire spathe surface or mainly near the midrib (vs. such cells completely lacking). The new species differs from both other species by its generally smaller stature (but see below) and seeds with numerous, knobby, transversely interrupted ridges as compared with generally fewer, smoother, uninterrupted ridges in the seeds of Polyspatha paniculata and testa shallowly ridged-reticulate in Polyspatha hirsuta (Figure 1: 7, 8; Plate 1).
Leaf pubescence in Polyspatha hirsuta can be quite variable, but plants with sparse pubescence or with long hairs confined to the adaxial leaf surface are restricted to Nigeria and further west, overlapping with Polyspatha oligospatha only in Ivory Coast. Thus throughout nearly all of the range of Polyspatha oligospatha, the pubescence differences of the leaf lamina, sheath, and internodes between Polyspatha oligospatha and Polyspatha hirsuta are consistent. The often much less pubescent plants of West African Polyspatha hirsuta still exhibit the stature, leaf shape and spathes that are typical of the species, so they would not be readily confused with Polyspatha oligospatha even if the two species were more broadly sympatric.
The new species has been confused in the past with small specimens of Polyspatha paniculata which can be of similar stature and appearance. Some may even have very similar looking spathes and smaller capsules and seeds than in typical plants of Polyspatha paniculata. Such plants can be separated from Polyspatha oligospatha by the absence of long, uniseriate hairs from the leaves, the spathes generally more crowded, overlapping and deflexed against the inflorescence axis, and the seeds with generally fewer, smooth, uninterrupted ridges, as in typical Polyspatha paniculata (Plate 1–1).
This species has been overlooked or misinterpreted partly because of Clarke’s Polyspatha paniculata var. glaucescens, which was raised to a species in the first edition of Flora of West Tropical Africa (FWTA) by
The collection Letouzey 11465 (Figure 2, arrow) from Cameroon is accepted as Polyspatha oligospatha with reservation. The pubescence of the upper leaf surface is more like that of Polyspatha hirsuta, with the uniseriate hairs longer, denser and paler than in typical Polyspatha oligospatha. However the absence of uniseriate hairs from the lower leaf surface, sheaths (except for the summits) and internodes distinguishes it from all Cameroonian specimens of Polyspatha hirsuta. Moreover, the outer surface of the small spathes is composed solely of lustrous, dark brown, bead-like cells, which are present in all specimens of Polyspatha oligospatha but none of Polyspatha hirsuta. The short marginal hairs on the spathes also agree with Polyspatha oligospatha, but are unusual for Polyspatha hirsuta in the same region. Overall, Letouzey 11465 is best treated as an atypical specimen of Polyspatha oligospatha.
Three sheets of Letouzey 11465 have been seen but those at P and YA were not critically examined. Thus my observations and conclusions have been based solely on the duplicate of this collection from BR. Unfortunately, it lacks seeds, which would be diagnostic.
Manning 2166 is unusual for Polyspatha oligospatha in several characters. The internodes and surface of the leaf sheaths bear numerous uniseriate hairs, a pubescence that has not been noted in any other collection of Polyspatha oligospatha. The spathes are also more pubescent than usual, bearing many, long, uniseriate hairs on the surface as well as along the midrib. The longest marginal hairs are >1.5 mm long, the normal maximum for Polyspatha oligospatha. Moreover, the spathes have very few lustrous, brown, bead-like cells and these are confined to the midrib area towards the spathe bases. In all these characters Manning 2166 resembles Polyspatha hirsuta. It differs from that species by its shorter stature, absence of long hairs from the abaxial leaf surface--they are typically present in Cameroonian collections--shorter uniseriate hairs on the internodes and sheath surfaces, and more spaced, patent and pointed spathes that bear at least a small number lustrous, brown, bead-like cells along the midrib. Manning 2166 might possibly represent a hybrid of Polyspatha oligospatha and Polyspatha hirsuta, but in view of the different times of anthesis in these species, I think it is best considered an atypical specimen of Polyspatha oligospatha. As in the case of Letouzey 11465, only the WAG sheet of Manning 2166, of the three specimens seen, was studied in great detail, thus reinterpretation might be necessary when the other sheets can be reexamined.
Polyspatha oligospatha has sometimes been collected as part of mixed collections with Polyspatha paniculata. I have seen five such mixed collections: Lieberberg 922 & 922A(K) from Uganda, Seret 70 (BR) from the Democratic Republic of Congo, Schlechter 12714 (BR, K) from the Republic of Congo, and Letouzey 8742 from Cameroon. Possibly when further duplicates of already seen collections are examined in additional institutions, other mixed collections will be discovered. Thus far no mixed collections have been seen of Polyspatha oligospatha and Polyspatha hirsuta.
The disjunct occurrence of Polyspatha oligospatha
in Ivory Coast is probably not an artifact of under-collection because
in both Ghana and Nigeria, the two larger of the four countries between
Ivory Coast and Cameroon, there were collectors who took a great
interest in Commelinaceae. J. K. Morton made numerous collections of Commelinaceae in Ghana and elsewhere and wrote several important papers on the family. D. P. Stanfield was an avid collector of Commelinaceae in Nigeria and was rewarded for his efforts and lengthy and detailed notes by having a new genus Stanfieldiella named for him by
Another factor that makes this disjunct distribution interesting is that it places Polyspatha oligospatha
on both sides of the Dahomey gap, the natural break in the rainforest
that separates the Upper Guinean and Congolian forest blocs. A similar
distribution pattern is shown by another Commelinaceae species, Palisota ambigua (P.Beauv.) C.B.Clarke, which Flora of West Tropical Africa (
Polyspatha oligospatha
appears to be reproductively isolated from the other two species
because of its morning anthesis and flowers fading about noon. In Flora
of West Tropical Africa
Polyspatha oligospatha Faden, a new species described herein, is a widespread African rainforest species that has been overlooked but is readily distinguished from the other two species by its seed testa pattern, leaf pubescence, spathe characters and morning flowering.
I thank Stanley Yankowski, with the assistance of Scott Whittaker, for the seed images; Alice Tangerini for the line drawings, organization of the seed images into Plate 1, and final modification of the distribution map; Sara Alexander for the distribution map; and the directors of BM, BR, EA, K, NY, MO, P, WAG and YA for access to their collections and/or loan(s) of specimens.